
Acute ventilatory responses to swimming at increasing intensities
- Published
- Accepted
- Received
- Academic Editor
- Davor Plavec
- Subject Areas
- Biophysics, Sports Medicine
- Keywords
- Ventilation, Respiratory frequency, Tidal volume, Breathing patterns, Swimming
- Copyright
- © 2023 Monteiro et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2023. Acute ventilatory responses to swimming at increasing intensities. PeerJ 11:e15042 https://doi.org/10.7717/peerj.15042
Abstract
Background
Physical exercise is a source of stress to the human body, triggering different ventilatory responses through different regulatory mechanisms and the aquatic environment imposes several restrictions to the swimmer, particularly regarding the restricted ventilation. Thus, we aimed to assess the acute ventilatory responses and to characterize the adopted breathing patterns when swimming front crawl at increasing intensity domains.
Methods
Eighteen well-trained swimmers performed 7 × 200 m front crawl (0.05 m∙s−1 velocity increments) and a maximal 100 m (30 s rest intervals). Pulmonary gas exchange and ventilation were continuously measured (breath-by-breath) and capillary blood samples for lactate concentration ([La−]) analysis were collected at rest, during intervals and at the end of the protocol, allowing the identification of the low, moderate, heavy, severe and extreme intensity domains.
Results
With the swimming velocity rise, respiratory frequency (fR), [La−] and stroke rate (SR) increased ([29.1–49.7] breaths∙min−1, [2.7–11.4] mmol∙L−1, [26.23–40.85] cycles; respectively) and stroke length (SL) decreased ([2.43–2.04] m∙min−1; respectively). Oxygen uptake (VO2), minute ventilation (VE), carbon dioxide production (VCO2) and heart rate (HR) increased until severe ([37.5–53.5] mL∙kg−1∙min−1, [55.8–96.3] L∙min−1, [32.2–51.5] mL∙kg−1∙min−1 and [152–182] bpm; respectively) and stabilized from severe to extreme (53.1 ± 8.4, mL∙kg−1∙min-1, 99.5 ± 19.1 L∙min−1, 49.7 ± 8.3 mL∙kg−1∙min−1 and 186 ± 11 bpm; respectively) while tidal volume (VT) was similar from low to severe ([2.02–2.18] L) and decreased at extreme intensities (2.08 ± 0.56 L). Lastly, the fR/SR ratio increased from low to heavy and decreased from severe to the extreme intensity domains (1.12 ± 0.24, 1.19 ± 0.25, 1.26 ± 0.26, 1.32 ± 0.26 and 1.23 ± 0.26).
Conclusions
Our findings confirm a different ventilatory response pattern at extreme intensities when compared to the usually evaluated exertions. This novel insight helps to understand and characterize the maximal efforts in swimming and reinforces the importance to include extreme efforts in future swimming evaluations.
Introduction
Breathing is a natural and fundamental human behavior that allows the exchange of respiratory gases between the lungs and the atmosphere. When we are under stress, as physical exercise, minute ventilation ( E) increases (Pelarigo et al., 2016; Tipton et al., 2017) due to the respiratory frequency (fR) and tidal volume (VT) rise. During an incremental exercise, fR increases non-linearly and VT tends to present a plateau, with the E rise at lower intensities depending on both fR and VT increases. The further growth in E at higher exercise intensities seems to be explained by the increase in fR, a phenomenon known as the tachypneic breathing pattern (Sheel & Romer, 2012; Nicolò, Marcora & Sacchetti, 2020). Despite the well-established knowledge on the fR and VT contributions for E increase during an incremental exercise, further research focusing on the different regulatory mechanisms that drive these contributions is welcome (Figueiredo et al., 2013; Tipton et al., 2017).
Central command, muscle afferent feedback and metabolic inputs are the major E behavior determinants despite acting with different timings when exercise intensity changes (Forster, Haouzi & Dempsey, 2012; Duffin, 2014; Tipton et al., 2017). The regulation of fR and VT is less studied but it was previously suggested that the inputs driving E act separately on these variables, with central command and muscle afferent feedback preferentially regulating fR (Amann et al., 2010; Nicolò et al., 2017), while metabolic responses are responsible for the VT regulation (Nicolò et al., 2017). Considering the great importance of the central command on fR control and the close association between breathing patterns, exercise modes and limbs movement (Sheel & Romer, 2012; Forster, Haouzi & Dempsey, 2012), it is of great importance to understand how the ventilatory response adapts to different exercise related constraints.
In swimming, the aquatic environment imposes significant restrictions on the human body, such as the increase of the hydrostatic pressure around the chest, resulting in an augmented work of the inspiratory muscles (Lomax & McConnell, 2003; Leahy et al., 2019). In addition, the swimming typical horizontal position leads to face immersion and, consequently, to restricted ventilation (Holmér et al., 1974; McCabe, Sanders & Psycharakis, 2015). These restrictions oblige swimmers to synchronize active inspiratory and expiratory phases with movements of upper and lower limbs, resulting in specific swimming breathing patterns (Leahy et al., 2019). Front crawl is the most common (in training and competition conditions) from the four swimming conventional techniques, with swimmers more generally inspiring on every two or three upper limbs actions, i.e., using unilateral and bilateral breathing patterns (Seifert, Chollet & Allard, 2005; Figueiredo et al., 2013). Despite the existing variability of the V˙E responses along the different intensity domains, particularly when using the front crawl technique (Ribeiro et al., 2015; Monteiro et al., 2022), the fR and VT behaviors are still scarcely studied when swimming at increasing paces.
Since further research on the fR and VT responses is necessary to improve the overall understanding of breathing physiology and ventilatory control, we have aimed to assess the acute ventilatory responses when swimming from low to extreme intensity domains. For achieving that purpose, swimmers were required to wear a breathing snorkel attached to a gas analyzer along a standard incremental front crawl protocol. Complementarily, we aimed to characterize the swimmers breathing patterns along the exercise intensity rise to understand if the synchronization with the upper and lower limbs motion is maintained even when using the respiratory snorkel, i.e., without constraining the inspiratory and expiratory phases. We have hypothesized that: (i) despite the respiratory constraints, gas exchange variables increase concomitantly with the swimming velocity rise, with fR and VT presenting a nonlinear increase and a stabilization (respectively); and (ii) swimmers keep the breathing patterns used in free swimming when breathing into a snorkel (due to the breathing synchronization with stroke rate).
Materials and Methods
Participants
Eighteen well-trained swimmers (nine males) volunteered to participate in the current study. Their main anthropometric, training background and competitive characteristics were (for males and females, respectively): 20.1 ± 8.0 vs. 16.8 ± 1.8 years of age, 176.6 ± 7.6 vs. 163.4 ± 4.7 cm of body height, 67.5 ± 12.1 vs. 57.3 ± 6.5 kg of body mass, 21.5 ± 2.7 vs. 21.4 ± 1.6 kg∙m−2 of body mass index, 8.3 ± 3.8 vs. 7.3 ± 3.4 years of swimming practice and 489 ± 66 vs. 478 ± 83 Fédération Internationale de Natation points of their best competitive performance event. Participants were recruited via personal contact and based on the following eligibility criteria: (i) without a history of cardiorespiratory and physical diseases or injuries within the previous 6 months; (ii) having ≥2 years of swimming training background and (iii) being engaged at ≥5 training sessions per week. All the experiments were approved by the Faculty of Sport of University of Porto ethics committee (CEFADE 25 2020) and participants were informed about the purpose, benefits and any associated risks (providing their written individual consent for participation in accordance with the Helsinki Declaration).
Experimental protocol
Subjects were asked to be rested and fully hydrated, and refrained from alcohol and caffeine consumption (and from vigorous exercise) for, at least, 24 h prior the evaluation. Test sessions were conducted in a 25 m indoor pool, with 27 °C and 26.5 °C of water and air temperatures (respectively) and 75% of humidity. After a 600 m low intensity in-water warm-up, each swimmer performed a front crawl discontinuous incremental protocol, consisting of 7 × 200 m (with 0.05 m∙s−1 velocity increments), followed by a maximal 100 m, with 30 s rest intervals in-between (adapted from Fernandes et al., 2005; Carvalho et al., 2020; Monteiro et al., 2022). The paces for each swimmer 7th step were established based on the individual 400 m front crawl performance on the evaluation day, then six velocity increments were subtracted. Swimming velocities were controlled using flashing lights on the bottom of the pool (Pacer2Swim; KulzerTEC, Aveiro, Portugal), with in-water starts and open turns (without underwater gliding) being used due to the impossibility of performing flip turns with deep water gliding when using a respiratory snorkel.
A portable gas analysis system (K4b2; Cosmed, Rome, Italy) was transported on a steel cable above the water surface allowing to measure breath-by-breath pulmonary gas exchange and ventilation by connecting the swimmer through a low hydrodynamic resistance respiratory snorkel and valve system (Aquatrainer®, Cosmed, Rome, Italy; Ribeiro et al., 2015). This gas analysis system was calibrated before each experimental session using ambient air against known concentrations (16% O2 and 5% CO2) and a 3 L calibration syringe. Heart rate (HR) was continuously recorded at the baseline and during the incremental protocol using a Polar Vantage NV (Polar Electro Oy, Kemple, Finland) that telemetrically emitted to the portable gas analyzer unit (de Jesus et al., 2015). Lactate concentration ([La−]) values were obtained using fingertip capillary blood samples collected at rest, immediately after the end of each step and at 1, 3, 5 and/or 7 min post-protocol (until obtaining maximal values) using a portable analyzer (Lactate Pro2; Arkay Inc., Kyoto, Japan; Carvalho et al., 2020). Stroke rate (SR) was assessed through the number of upper limbs cycles per minute in the last 50 m of each step (using a Finis stopwatch with a frequency meter function) and stroke length (SL) was calculated by dividing the mean velocity by SR (Fernandes et al., 2005; Monteiro et al., 2022).
Data analysis
The pulmonary gas exchange and ventilation data were examined to exclude occasional errant breaths (eventually caused by swallowing, coughing or signal interruptions). For the analysis, only the values of oxygen uptake ( O2) between ±3 SD were considered (Monteiro et al., 2020), which were then smoothed using a three breaths moving average and 10 s time average (Fernandes et al., 2012). The mean values from the last 30 s of exercise per step were selected and conventional physiological criteria were applied to stablish the maximal oxygen uptake O2max; (Howley, Bassett & Welch, 1995; Zacca et al., 2020). The lactate-velocity curve modelling method, through the determination of the interception point of the best fit of a combined linear and exponential pair of regressions, was used to determine the individual anaerobic threshold (Carvalho et al., 2020; Monteiro et al., 2022). Using the O2max and the anaerobic threshold as physiological indicators, the following intensity domains were identified (Fig. 1): (i) the low and moderate domains, corresponding to two steps below and the step at the anaerobic threshold; (ii) the heavy and severe domains, matching the step below and the step where O2max was elicited; and (iii) the extreme domain, allocated to the maximum 100 m at the end of the incremental protocol (Fernandes et al., 2012; de Jesus et al., 2015; Ribeiro et al., 2017). Swimmers breathing patterns were determined by calculating the ratio between fR and SR.
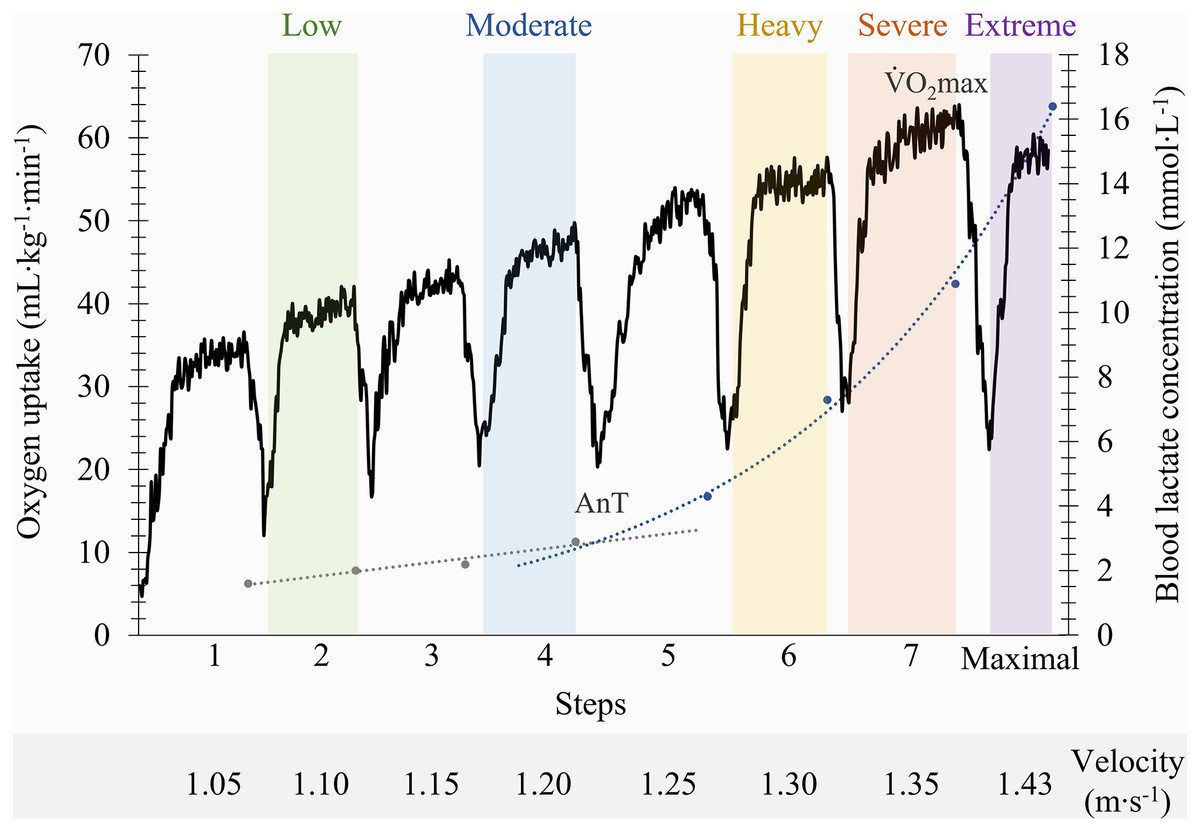
Figure 1: Identification of the front crawl swimming intensity domains on a representative participant based on anaerobic threshold (AnT) and maximal oxygen uptake (V˙O2max) indicators.
{kind=link}
Statistical analysis
A sample size of 18 subjects was required for a paired sample design to detect a moderately large effect size (0.83) with a 5% significance level and 95% power (G*Power 3.1.9.7; Heinrich Heine Universität Düsseldorf, Düsseldorf, Germany). Statistical procedures were conducted using SPSS (version 27.0.1.0; IBM Corporation, Armonk, NY, USA) and the normal data distribution was checked for all variables using the Shapiro-Wilk test. Mean and standard deviation values were computed for descriptive analysis of all variables and one-way repeated measures analysis of variance with Bonferroni adjustment was used to compare the assessed physiological and performance variables along the intensity domains spectrum (p ≤ 0.05 level). Partial eta-squared ( ) for effect size calculation was computed to compare the magnitude of changes between swimming intensity domains.
Results
Depending on the swimmer, the establishment of the low and moderate intensity domains corresponded to the swimming velocity between the first-third and third-fifth steps (respectively), while the heavy, severe and extreme intensity domains corresponded to the sixth, seventh and maximal last protocol steps (in this order). The low, moderate, heavy, severe and extreme efforts were performed at 1.04 ± 0.11, 1.13 ± 0.11, 1.22 ± 0.10, 1.26 ± 0.10 and 1.39 ± 0.11 m∙s−1 (respectively) and all the physiological and performance variables are presented in Fig. 2. With the swimming intensity rise (Z4.68 = 305.79, = 0.95, p < 0.001), fR, [La−] and SR increased ( = 0.81, = 0.95 and = 0.88, respectively; p < 0.001) and SL decreased ( = 0.59, p < 0.001). O2, E, CO2 and HR increased from low to severe intensities ( = 0.91, = 0.88, = 0.89 and = 0.83, respectively; p < 0.001), but all stabilized at extreme exertion and VT presented similar values from low to severe exertions and decreased at the extreme intensity (p = 0.006). The fR/SR ratio increased from low to moderate (p = 0.01) and from moderate to heavy domains (p = 0.02) and lower values were observed at extreme compared to severe intensity (p = 0.02; Fig. 3).
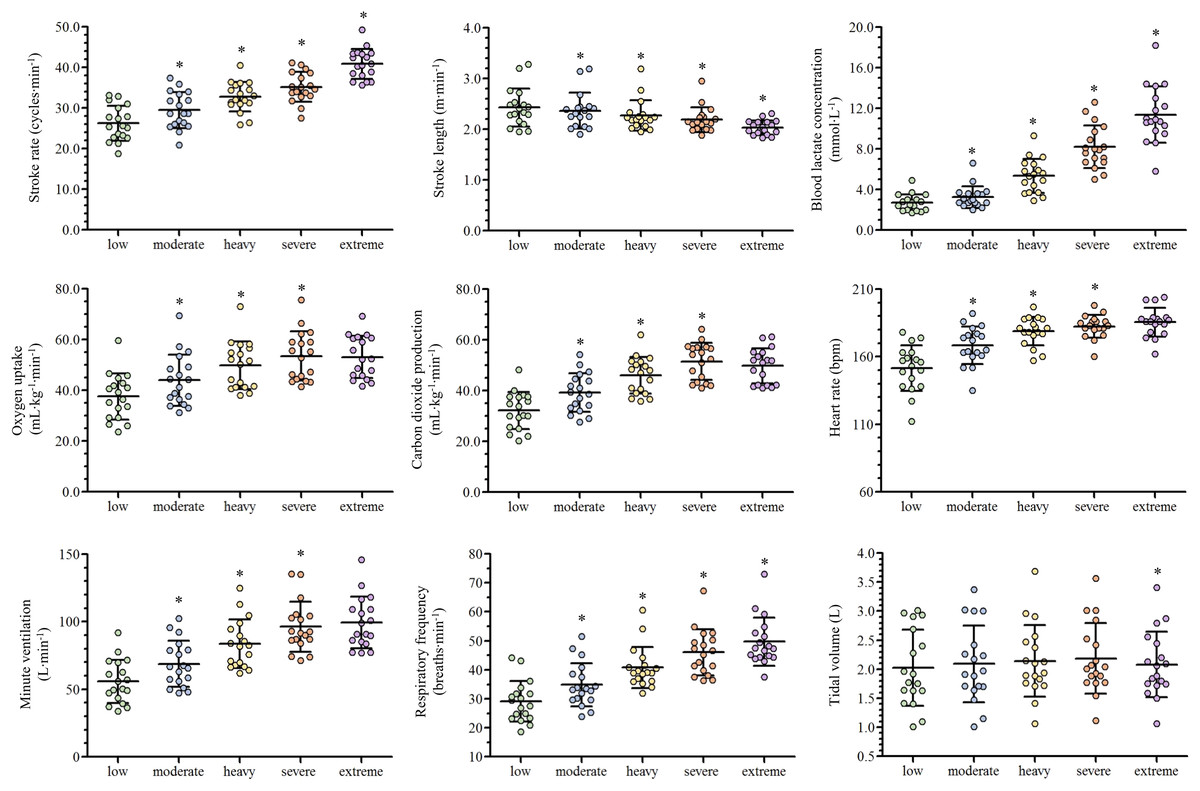
Figure 2: Scatter plots with mean and standard deviation of the ventilatory and performance variables responses along with the swimming intensity rise.
An asterisk (*) indicates difference from the previous intensity domain (p < 0.05).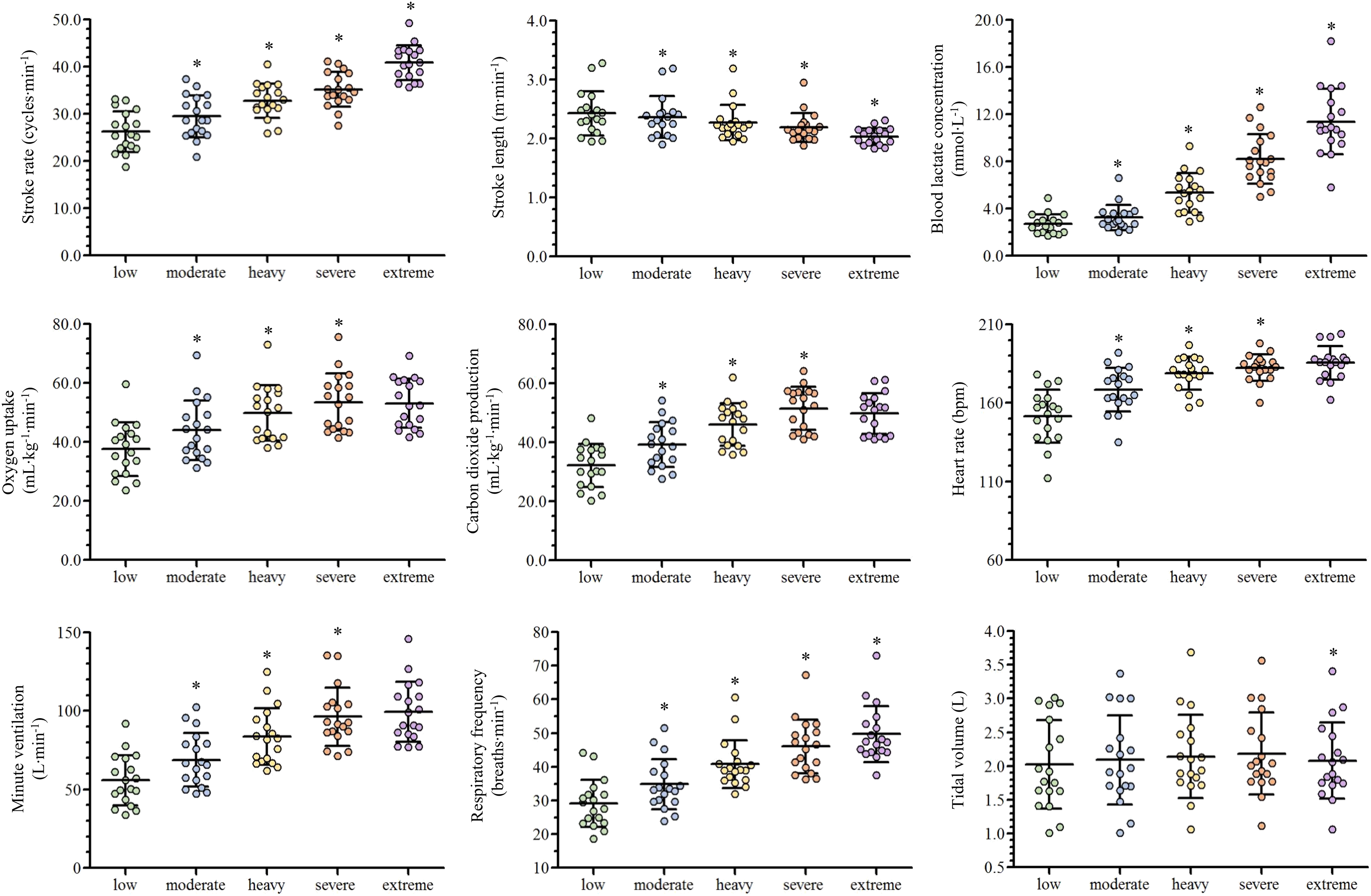{kind=link}
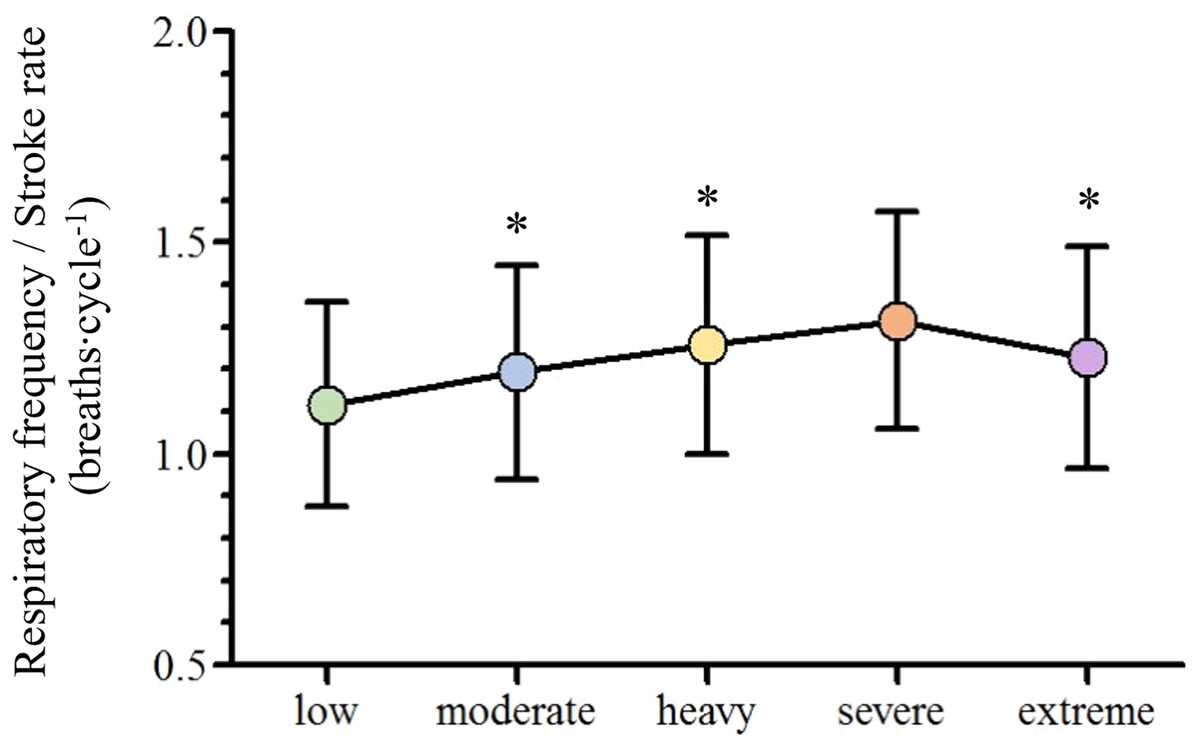
Figure 3: Mean and standard deviation of the adopted breathing patterns along with the swimming intensity rise.
An asterisk (*) indicates difference from the previous intensity domain (p < 0.05).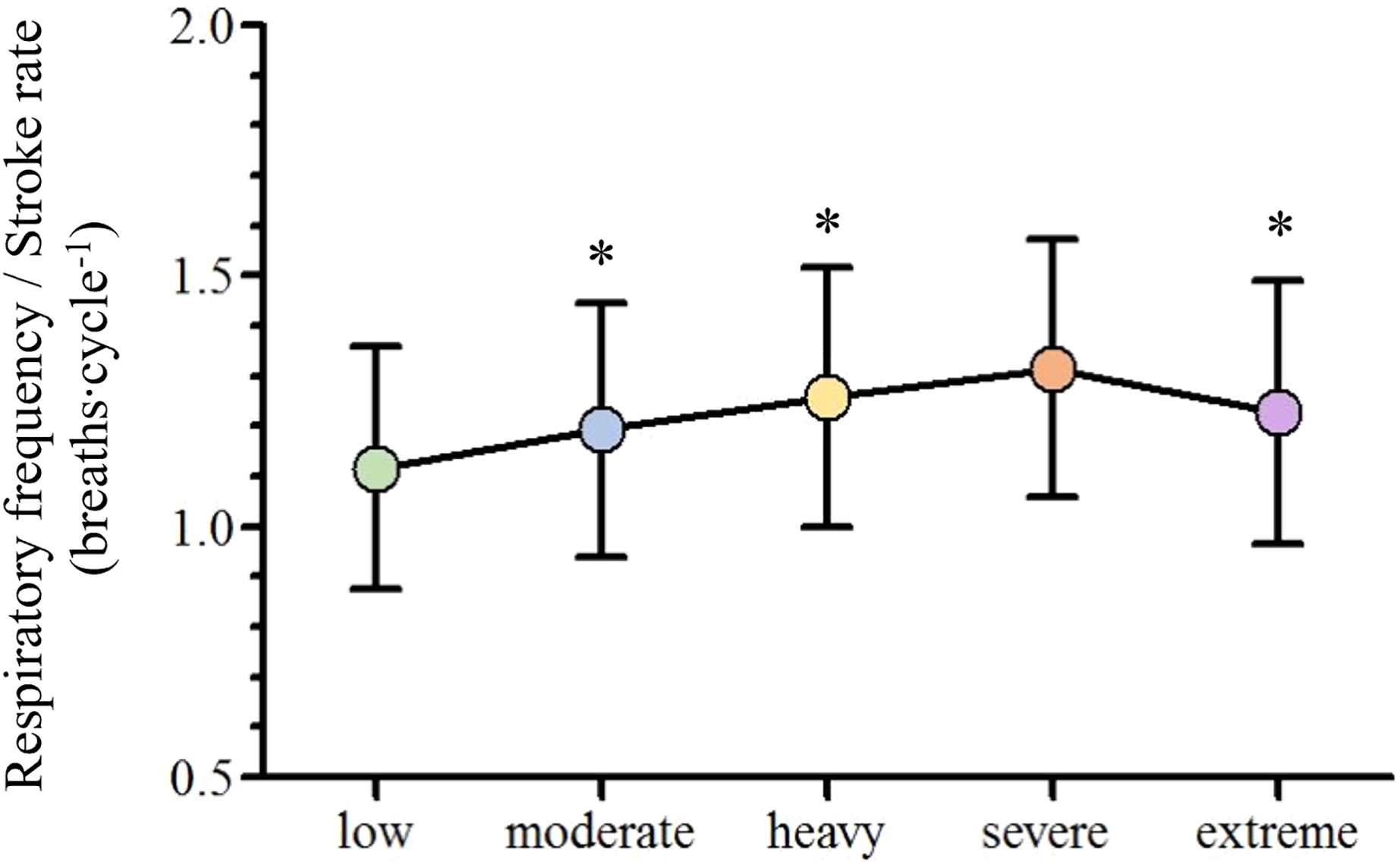{kind=link}
Discussion
The main purpose of the current study was to assess swimmers acute ventilatory responses when performing front crawl from low to extreme intensities. As hypothesized, the values of the selected gas exchange variables increased along with the swimming intensity rise (until the severe intensity domain) and fR and VT presented a nonlinear increase and a plateau (respectively) in the incremental protocol. Concurrently, we aimed to analyze swimmers breathing pattern behavior as swimming pace was rising, being observed an fR/SR ratio increase until reaching the severe intensity domain and a decrease at extreme exertions. This does not confirm our initial hypothesis that the front crawl breathing pattern was going to remain stable when swimming with a respiratory snorkel.
It is well established that the 7 × 200 m front crawl intermittent incremental protocol allows collecting capillary blood for [La−] analysis (Fernandes et al., 2005; Pelarigo et al., 2016; Carvalho et al., 2020) and, together with the gas exchange assessment, ensures a complete physiological characterization of the low, moderate, heavy and severe intensity domains (de Jesus et al., 2015; Zacca et al., 2019; Monteiro et al., 2022). The current results are similar to those previously presented for the low-severe swimming intensity domains when using the same methodological approach, particularly regarding V˙O2 (Fernandes et al., 2012; de Jesus et al., 2015), E and fR (Pelarigo et al., 2016; Monteiro et al., 2022), and [La−] values (Štrumbelj et al., 2007; Sousa, Vilas-Boas & Fernandes, 2014; Monteiro et al., 2022).
However, most official swimming events (such as the 50, 100 and 200 m distances) occur at the extreme intensity domain, and is why a complete swimming ventilatory characterization should also include extreme efforts. This is a fundamental training zone for excelling competitive swimmers performances where the exertions are so intense that fatigue occurs and exercise ends before O2max can be reached (Hill, Poole & Smith, 2002; Ribeiro et al., 2017). Studies focusing on the anaerobic capacity development are very scarce, with swimmers acute ventilatory responses remaining almost unexplored, justifying the inclusion of a 100 m maximal bout at the end of the front crawl incremental protocol. This maximal intensity short duration effort, when swimming up at the standard 7 × 200 m step protocol, allows swimmers to have their physiological profile fully characterized.
The current ventilatory results at the maximal 100 m front crawl evidenced lower O2 and V˙E values compared to those reported for the same intensity domain (Sousa, Vilas-Boas & Fernandes, 2014), probably due to the sample characteristics (higher level and male swimmers only) and the higher swimming velocity achieved (the current 100 m bout was part of an incremental protocol instead of an isolated rectangular test). Additionally, higher fR values were observed at the current extreme intensity domain compared to a maximal 200 m front crawl bout performed at a lower swimming velocity (Štrumbelj et al., 2007). This demonstrates that the extreme exertions (only inferiorly delimited by the O2max) can include a wider range of swimming velocities, being important to consider them when comparing the results obtained at this intensity domain.
Swimming faster implied a O2, V˙E, fR, CO2, [La-] and SR increase and a SL decrease from low to severe exertions, as previously described (Figueiredo et al., 2013; de Jesus et al., 2015; Monteiro et al., 2022), while O2, V˙E, CO2 and HR stabilized from severe to extreme intensities (Sousa, Vilas-Boas & Fernandes, 2014). The attainment of O2max at severe intensity paces, and the fact that these abovementioned variables are highly related, explain the maintenance of similar values despite the swimming intensity rise (Sousa, Vilas-Boas & Fernandes, 2014; Nicolò et al., 2018; Monteiro et al., 2022). HR at O2max corresponded to 90.4% ± 3.1% of its maximum, in accordance with the secondary criteria used to confirm O2max (Howley, Bassett & Welch, 1995; Zacca et al., 2020). Maximal [La−] values were observed at extreme exertions, where the energy production is highly dependent on anaerobic metabolism, with a higher production of lactate and, consequently, its progressive accumulation in the bloodstream (Hargreaves & Spriet, 2020). Contrarily to what was described (Sousa, Vilas-Boas & Fernandes, 2014), our [La−] values increase from severe to extreme intensity domains is explained by a bigger velocity rise (~10% instead of 5%), corroborating the existence of a wide range of swimming velocities at this domain.
The observed V˙E, fR and VT behaviors along the low to extreme swimming intensity domains spectrum corroborates what is described in the literature, independently of the exercise modality performed (Amann et al., 2010; Nicolò et al., 2017). This seems to indicate that, regardless of the swimming movements and the different constraints imposed by the aquatic environment, the central command, muscle afferent feedback and metabolic inputs have the same influence on their regulation along the intensity domains spectrum (Štrumbelj et al., 2007; Sheel & Romer, 2012; Forster, Haouzi & Dempsey, 2012). However, the swimming intensity rise resulted in the selection of different breathing patterns at each intensity domain. Diversely to what was initially expected, the fR/SR ratio tended to increase until the heavy intensity domain, indicating that swimmers took advantage of free breathing while using the respiratory snorkel (Štrumbelj et al., 2007). The fR/SR ratio decrease from severe to extreme intensity domains can be justified by both the maximal intensity and the short time duration effort of the 100 m exertion, where SR increased more than fR (16% vs. 8%, respectively). In addition, the lower increase in fR compared to SR seems to indicate that this extreme effort is characterized by moments of apnea.
Conclusions
The fR and, consequently, the fR/SR ratio values were influenced by the use of the respiratory snorkel and its interpretation may be different compared to free swimming. However, this is the only methodology that provides a real time and breath-by-breath assessment of the swimmers ventilatory responses. In conclusion, by proposing the addition of a maximal effort at the end of the front crawl intermittent incremental swimming protocol, the current study provides a novel framework of the acute ventilatory responses to the large spectrum of swimming intensity domains, particularly at the extreme exertion, used both in training and competition contexts. V˙O2, V˙E and V˙CO2 stabilized, and VT decreased, from severe to extreme intensity domains, differently to what happened from low to severe exertions, while fR and SR increased along the swimming intensities spectrum. The breathing pattern varied along the incremental protocol and its synchronization with stroke rate was not verified when using the respiratory snorkel.
Supplemental Information
Raw ventilatory and performance data.
Ventilatory and performance variable responses along the swimming intensity domains.