
Whole-genome analysis of Escherichia coli isolated from wild Amur tiger (Panthera tigris altaica) and North China leopard (Panthera pardus japonensis)
- Published
- Accepted
- Received
- Academic Editor
- Bernd Neumann
- Subject Areas
- Bioinformatics, Genomics, Microbiology, Veterinary Medicine, Zoology
- Keywords
- Escherichia coli, Amur tiger, North China leopard, Whole-genome sequencing, Drug resistance
- Copyright
- © 2024 Li et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2024. Whole-genome analysis of Escherichia coli isolated from wild Amur tiger (Panthera tigris altaica) and North China leopard (Panthera pardus japonensis) PeerJ 12:e17381 https://doi.org/10.7717/peerj.17381
Abstract
Background
Escherichia coli is an important intestinal flora, of which pathogenic E. coli is capable of causing many enteric and extra-intestinal diseases. Antibiotics are essential for the treatment of bacterial infections caused by pathogenic E. coli; however, with the widespread use of antibiotics, drug resistance in E. coli has become particularly serious, posing a global threat to human, animal, and environmental health. While the drug resistance and pathogenicity of E. coli carried by tigers and leopards in captivity have been studied intensively in recent years, there is an extreme lack of information on E. coli in these top predators in the wild environment.
Methods
Whole genome sequencing data of 32 E. coli strains collected from the feces of wild Amur tiger (Panthera tigris altaica, n = 24) and North China leopard (Panthera pardus japonensis, n = 8) were analyzed in this article. The multi-locus sequence types, serotypes, virulence and resistance genotypes, plasmid replicon types, and core genomic SNPs phylogeny of these isolates were studied. Additionally, antimicrobial susceptibility testing (AST) was performed on these E. coli isolates.
Results
Among the E. coli isolates studied, 18 different sequence types were identified, with ST939 (21.9%), ST10 (15.6%), and ST3246 (9.4%) being the most prevalent. A total of 111 virulence genes were detected, averaging about 54 virulence genes per sample. They contribute to invasion, adherence, immune evasion, efflux pump, toxin, motility, stress adaption, and other virulence-related functions of E. coli. Sixty-eight AMR genes and point mutations were identified. Among the detected resistance genes, those belonging to the efflux pump family were the most abundant. Thirty-two E. coli isolates showed the highest rate of resistance to tetracycline (14/32; 43.8%), followed by imipenem (4/32; 12.5%), ciprofloxacin (3/32; 9.4%), doxycycline (2/32; 6.3%), and norfloxacin (1/32; 3.1%).
Conclusions
Our results suggest that E. coli isolates carried by wild Amur tigers and North China leopards have potential pathogenicity and drug resistance.
Introduction
Escherichia coli represents a major cause of morbidity and mortality worldwide (Da Silva & Mendonça, 2012). Essentially, E. coli is sensitive to almost all clinically relevant antimicrobial agents; however, with the widespread use of antibiotics, the resistance rate of E. coli to a variety of drugs has gradually increased, and the infections caused by its drug-resistant strains have tended to increase, and its drug-resistance has continued to transform (Liu et al., 2021). Over the past decades, an increasing number of resistance genes have been identified in E. coli isolates, and these resistance genes have been transferred horizontally enabling E. coli to acquire resistance, e.g., through resistance plasmids, or other mobile genetic elements, transposons, and gene cassettes in class I and class II integrons (Poirel et al., 2018).
One of the biggest obstacles to control and treat infectious diseases in animals is the resistance of the causative organisms to antibiotics (Duangurai et al., 2022). As antibiotic resistance increases in wildlife populations, previous antibiotic treatments are becoming ineffective, and fighting infectious diseases will become increasingly challenging (Lagerstrom & Hadly, 2021). While the drug resistance and pathogenicity of E. coli carried by tigers and leopards in captivity have been studied intensively in recent years, there is an extreme lack of information on E. coli in these top predators in the wild environment. By studying E. coli drug resistance in wild tigers and leopards, we can indirectly assess the spread of E. coli drug resistance genes in animal microcosms under the selective pressure of antibiotic use by humans, thus reflecting the ecological condition of wild tigers and leopards.
Several medications are used in traditional antimicrobial therapy to target the pathogen’s various functions. However, with every new drug and antibiotic used worldwide, bacteria continue to evolve new mechanisms to evade this drug-mediated killing at an alarming rate, a phenomenon known as Antimicrobial Resistance (AMR). Interest in AMR bacteria and AMR genes isolated from wildlife and the environment has recently grown. Several wildlife species have been found to harbor bacteria resistant to antimicrobial (Costa et al., 2008; Dolejska, Cizek & Literak, 2007; Poeta et al., 2005), and wildlife has been identified as a possible source of AMR bacteria and AMR genes (Arnold, Williams & Bennett, 2016). Wild Amur tigers and North China leopards, as rare and large wild animals in China, play a pivotal role in maintaining biodiversity and ecological balance within their habitats. However, the emergence of antibiotic-resistant E. coli in these endangered wild animals could pose a threat to their health and the ecosystems they inhabit. The antibiotic resistance genes of E. coli can spread through water sources, the food chain, or contact with humans and other wildlife (Koutsoumanis et al., 2021), leading to other individuals or species in the wildlife population becoming resistant as well, thus exacerbating the problem of antibiotic resistance and increasing the risk of spreading antibiotic-resistant pathogens in humans and other animals. The spread of drug-resistant E. coli in wildlife could disrupt the balance of ecosystems, where potentially pathogenic and resistant organisms from any one of these ecosystems could easily move to another ecosystem (Collignon & McEwen, 2019), leading to over-infection of some species, which could have indirect effects on other organisms, affecting multiple links in the ecological chain, including the food chain and natural interactions, and thus potentially posing threats.
In recent years, with the rapid development of genomics, whole genome sequencing technology has shown its unique advantages in species identification, drug resistance, virulence prediction, and genetic evolutionary analysis (Purushothaman, Meola & Egli, 2022). The distribution of bacterial resistance and virulence genes discovered through whole genome sequencing is then used to infer potential bacterial drug resistance and virulence phenotypes, which is crucial for the prevention and treatment of bacterial illnesses. Here, we performed whole-genome sequencing and comparative analysis of 32 E. coli isolates from the feces of wild Amur tiger and North China leopard.
Materials and Methods
Bacterial isolation
From 2012 to 2015, twenty-four fecal samples (H1–H24) from the Amur tiger were randomly collected with the permission of the management authorities of The Changbai Mountain Area in Jilin Province and the Wandashan Mountain National Nature Reserve in eastern Heilongjiang Province, including five samples in 2012, nine in 2013, six in 2014 and four in 2015, total 24. In November 2020, eight fecal samples (B1–B8) of North China leopards were randomly collected from the Liupan Mountains in southern Ningxia, China (Table S1). Fecal samples were transported to the laboratory after aseptic collection and stored at −80 °C for the next step in the experiment. The sampler did not introduce any harmful substances when collecting the feces, which would not disturb the animal’s habitat.
Isolation and identification of E. coli and whole genome sequencing
A sterile cotton swab was inserted into the middle of the fecal sample, dipped into an appropriate amount of feces, and spread evenly on MacConkey agar, and then placed into a 37 °C constant temperature incubator for 12 h, and pink single colonies could be observed. Then use the inoculation loop to inoculate the bacteria onto Eosin Methylene Blue agar until single colonies with black purple color and metallic luster grow, and then inoculate onto Nutrient Agar and incubate at 37 °C for 12 h until white single colonies grow.
The white single colonies were picked and tested using the Tubes for biochemical identification of Enterobacteriaceae (Hopebio, Qingdao, China). Subsequently, the genomic DNA of the strain was extracted using a genomic DNA purification kit (Tiangen, Beijing, China) and amplified by PCR with universal primers for bacterial 16S rRNA gene, and the 16S rRNA gene fragment of the strain was amplified and sent to Jilin Comate Bioscience Ltd., Jilin, China. After confirming the bacteria as E. coli based on the sequencing results and biochemical identification, subsequent experiments were performed.
Before library preparation, the DNA was fragmented and screened, and then the MGIEasy Universal DNA Library Preparation Reagent Kit (MGI, Shenzhen, China) was used for library preparation. In the process of library preparation, the DNA fragments were firstly end-repaired and dA tails were added, followed by junction ligation and purification of ligation products. Next, PCR amplification, PCR product purification, PCR product quality control, denaturation, and single-stranded cyclization were performed, followed by enzymatic digestion, enzymatic digestion product purification, and enzymatic digestion product quality control. The digested products were then sequenced on an MGISEQ-2000 gene sequencer from BGI Shenzhen, China, at 150 bp bipartite.
Antimicrobial susceptibility testing
Antimicrobial susceptibility testing was performed using the WHO-recommended Kirby-Bauer drug-sensitive paper diffusion method according to the standards of the Clinical and Laboratory Standards Institute (CLSI) (Díez-Aguilar et al., 2015). We selected five drug-sensitive papers from three classes, including fluoroquinolone antibiotics (ciprofloxacin, norfloxacin), tetracycline antibiotics (tetracycline, doxycycline), and penicillin antibiotics (imipenem). The selection of these drugs was based on resistance phenotypes corresponding to predicted resistant genotypes, and antibiotics of medical and veterinary concern were also considered.
Reads processing, assembly, and annotation
To ensure the reliability of subsequent credit analysis results, the quality of sequencing raw reads needs to be assessed first, and low-quality data needs to be removed. Here FastQC 0.11.9 (available online at https://www.bioinformatics.babraham.ac.uk/projects/fastqc/, accessed on 2 July 2023) was used to quality control the original sequencing reads. Then the data were filtered using fastp 0.23.2 (Chen et al., 2018) to remove adaptors, primers, and low-quality reads. To obtain a series of contigs for subsequent analysis, the sequence splicing of the Clean Data obtained after fasp filtering was performed using SPAdes 3.15.4 (Bankevich et al., 2012), where the parameter k-mer size list was set to 55, 65, 75, and 95.
The sequence splicing results obtained from the assembly were then statistically analyzed using Quast 5.2.0 (Gurevich et al., 2013) and Busco 5.6.1 (Seppey, Manni & Zdobnov, 2019). The analysis aimed to assess the integrity of the genome assembly.
Bioinformatics analysis
The assembled sequences of E. coli isolates were analyzed at the Center for Genomic Epidemiology (available online at https://www.genomicepidemiology.org/services/index.html, accessed on 15 February 2023) to identify multifocal sequence types, serotypes, plasmid replicons, virulence genes, and evolutionary relationships, respectively. Multi-locus sequence typing was performed by running MLST 2.0 (Larsen et al., 2012) based on the Achtman scheme for E. coli. Serotype identification of assembled sequences by SerotypeFinder 2.0 (Joensen et al., 2015) with a selected threshold of 90% identity and 60% total serotype gene length. PlasmidFinder 2.1 (Carattoli & Hasman, 2020) was used to study plasmid replicon sequences with thresholds of 90% identity and 60% minimum length, respectively. Abricate 0.5 with the virulence factor database (VFDB) (Chen et al., 2016) was used for identifying virulence genes, with parameters set to default values. The whole genome of E. coli MG1655 (NC_000913.3) was used as the reference, and assemblies of the 32 strains included in this study were uploaded to the CSI Phylogeny service to construct a phylogeny based on the concatenated alignment of high-quality SNPs. Whole genome data of some ST939 and ST10 strains were downloaded from NCBI for in-depth phylogenetic analysis with sequenced samples from this study. Parsnp 2.0.3 (Kille et al., 2024) was used to identify single nucleotide polymorphisms (SNPs) in the E. coli genome set and to estimate a phylogenetic tree based on these SNPs. The evolutionary tree was constructed using RAxML 8.2.13 (Stamatakis, 2014) with the maximum likelihood method and the bootstrap replication value was set to 1,000.
Assembly files generated by SPAdes 3.15.4 are used as input files to ResFinder 4.1 (Bortolaia et al., 2020) and CARD (Alcock et al., 2023) to identify both genes and point mutations that confer antimicrobial resistance.
Results
Antimicrobial susceptibility testing
In the present study, it was found that among the 32 E. coli isolates, there were some differences in the rate of resistance to different drugs through antimicrobial susceptibility testing. Specifically, the highest resistance rate to tetracycline was (14/32; 43.8%), followed by imipenem (4/32; 12.5%), ciprofloxacin (3/32; 9.4%), doxycycline (2/32; 6.3%) and norfloxacin (1/32; 3.1%) (Table 1). Notably, one sample showed resistance to all five antimicrobials (Table S2).
| Class of drug | Antimicrobial susceptibility testing | ||
|---|---|---|---|
| Resistant number | Intermediate number | Sensitive number | |
| Tetracycline | 14 | 3 | 15 |
| Imipenem | 4 | 10 | 18 |
| Ciprofloxacin | 3 | 3 | 26 |
| Doxycycline | 2 | 2 | 28 |
| Norfloxacin | 1 | 1 | 30 |
Genomic characteristics of Escherichia coli
Thirty-two strains of E. coli isolated from fecal samples of wild Amur tigers (Panthera tigris altaica, n = 24) and North China leopards (Panthera pardus japonensis, n = 8) were subjected to whole-genome sequencing. The original reads were assembled from scratch to produce overlapping clusters ranging in size from 4.50 Mbp to 5.40 Mbp, with an overall GC content between 50.17% and 50.93%. The number of coding sequences ranged from 4,246 to 5,186, and all samples had a 100% complete BUSCO rate. Consequently, the quality of all assembled overlapping clusters of the 32 E. coli isolates was deemed sufficient for genome-wide analysis. The number of coding sequences ranged from 4,246 to 5,186, thus the quality of all assembled overlap clusters of the 32 E. coli isolates was considered sufficient for genome-wide analysis. The general characteristics of the E. coli genome can be found in Table S3.
MLST, serotypes and plasmid characterization
Among the E. coli isolates studied, 18 different sequence types were identified, with ST939 (21.9%), ST10 (15.6%), and ST3246 (9.4%) being the most prevalent. Other clones were also observed: n = 2 each from ST58, ST126, and singletons from ST11910, ST4038, ST164, ST1611, ST2079, ST1377, ST5418, ST7188, ST1378, ST2715, ST1380, ST69 and ST422 (Fig. 1).
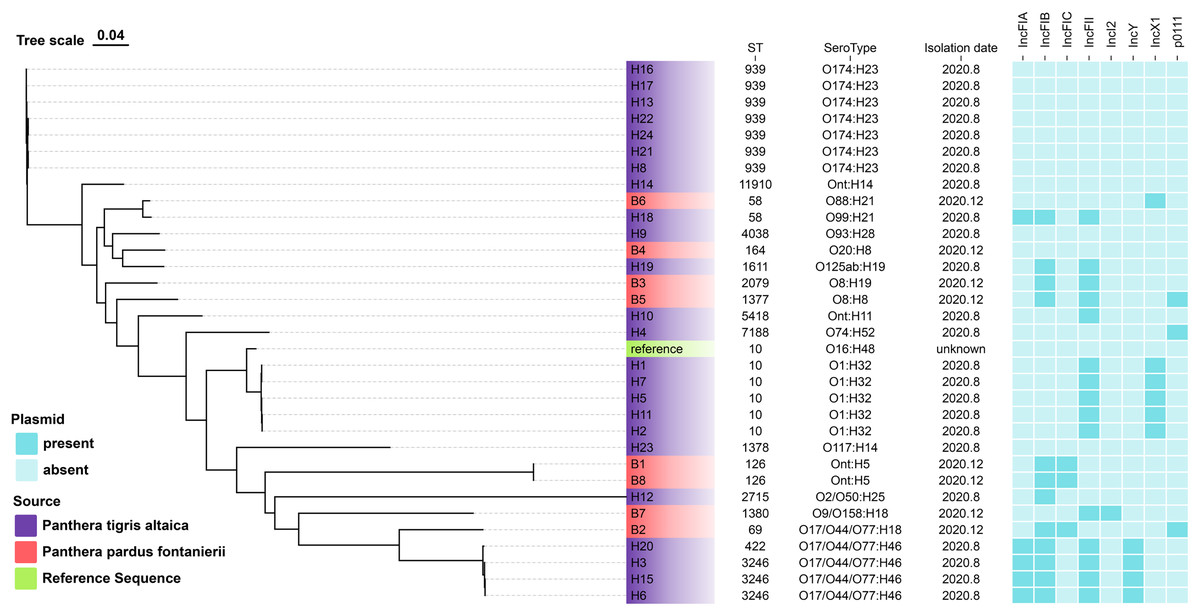
Figure 1: The evolutionary relationship of core genome-SNPs in the 32 E. coli isolates from Panthera pardus fontanierii and Panthera tigris altaica.
The phylogenetic tree was constructed using the core SNPs identified from 32 E. coli genome sequences and an E. coli reference genome downloaded from NCBI. The analysis was performed using CSI Phylogeny 1.4 (https://cge.food.dtu.dk/services/CSIPhylogeny/) with the Maximum Likelihood method and a default bootstrap replication value of 1,000. The E. coli phylogeny indicates (from left to right) sequence type (ST), serotype, isolation date, and plasmid replicon type for each strain.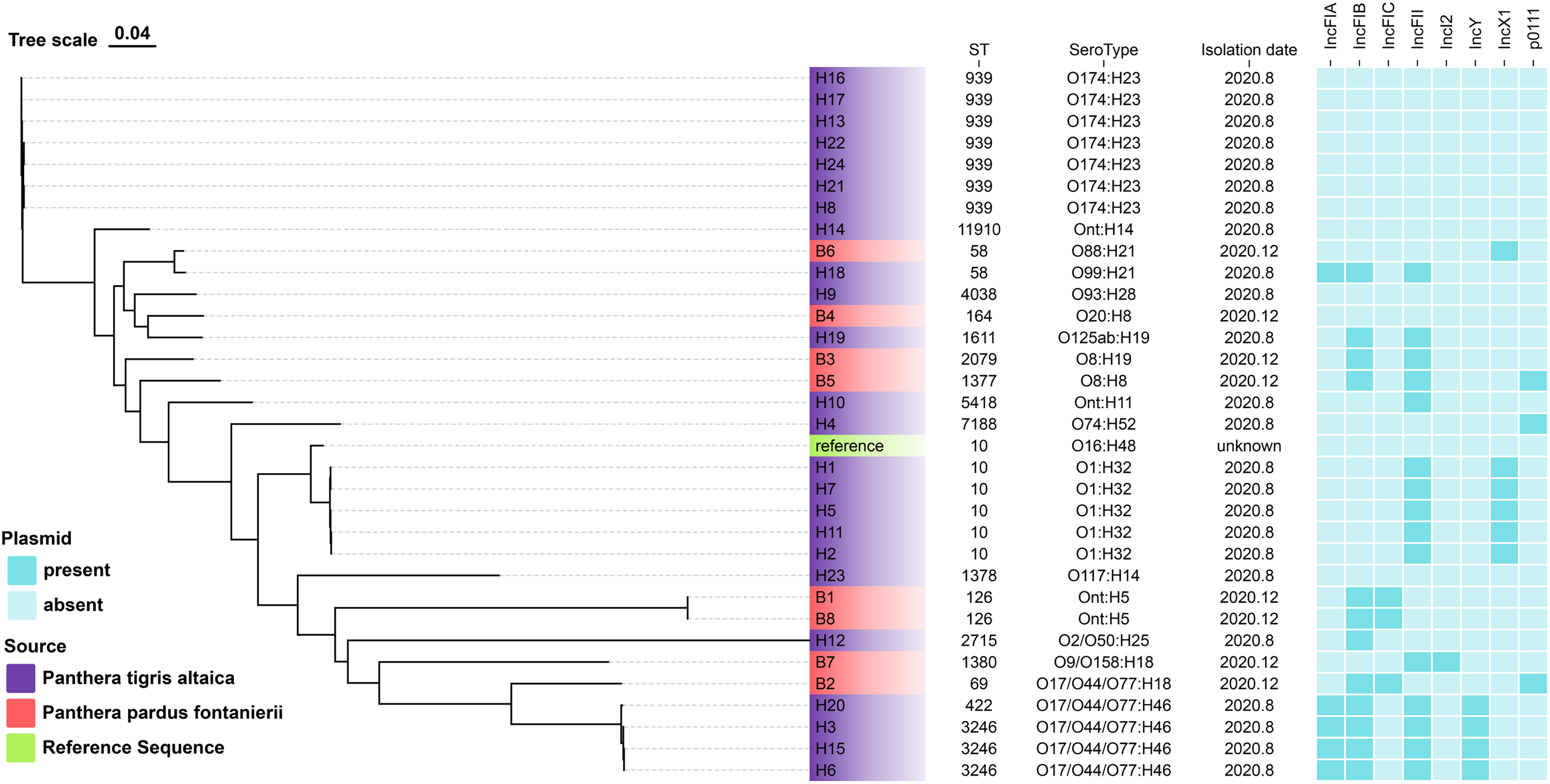{kind=link}
In silico plasmid replicon typing revealed that the IncF-type (59.4%) plasmid was the most common among the isolates included in this study. Other replicon types included IncX1, IncY, p0111, and IncI2. Among the IncF-type plasmids, IncFII (46.9%) was the most prevalent, followed by IncFIB, IncFIA, and IncFIC (Fig. 1).
We identified multiple serotypes in a collection of 32 E. coli isolates. The most predominant serotype was O174:H23 (21.9%), followed by O1:H32 (15.6%). Four isolates were founding the serogroups O17/O44/O77:H46. Two isolates were found in the serogroups Ont: H5. The detection rates of serum groups Ont:H14, O88:H21, O99:H21, O93:H28, O20:H8, O125ab:H19, O8:H19, O8:H8, Ont:H11, O74:H52, O16:H48, O117:H14, O2/O50:H25, O9/O158:H18, and O17/O44/O77:H18 were the lowest (Fig. 1).
Distribution of virulence factors and antibiotic-resistance genes
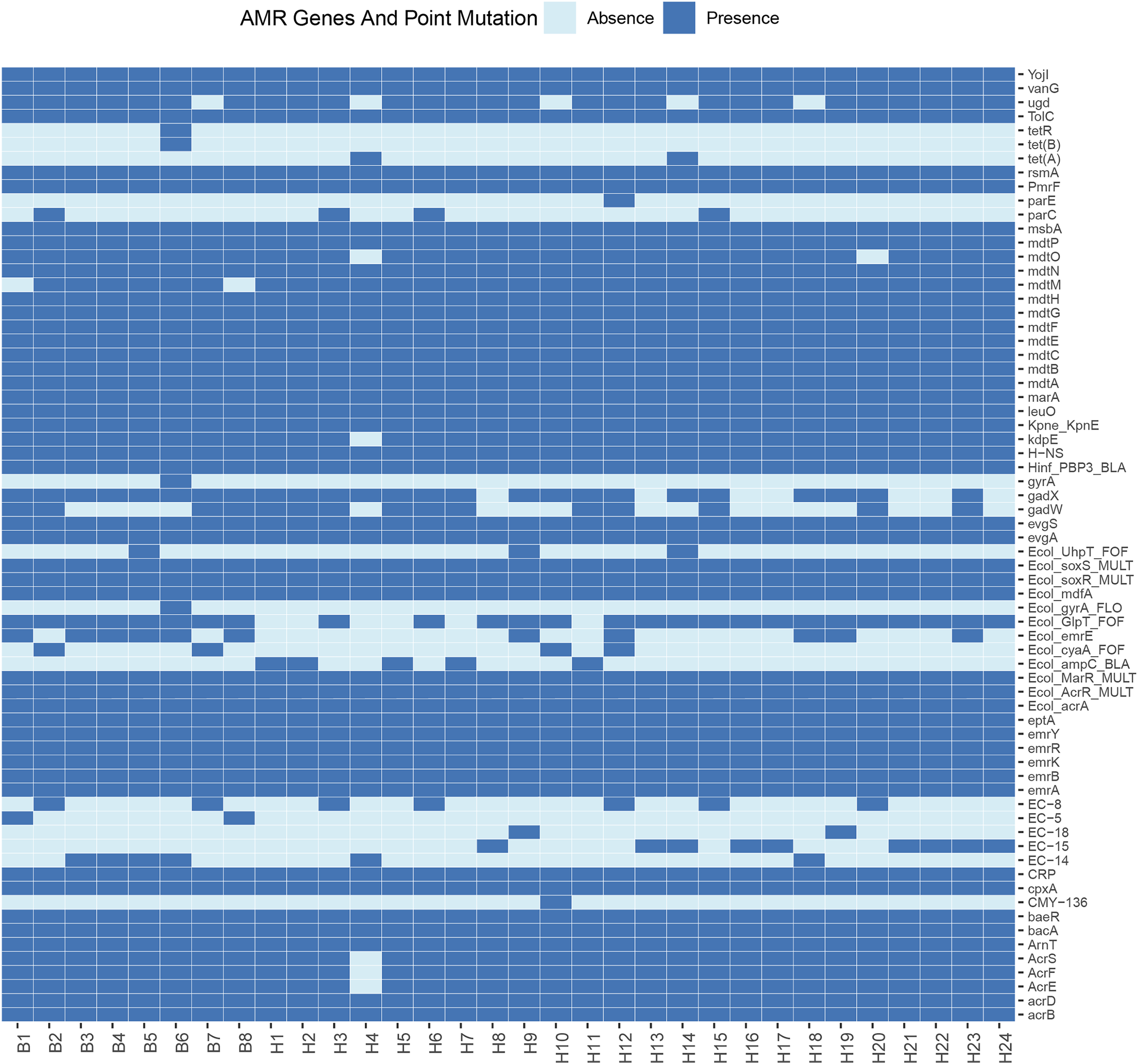
Sixty-eight resistance genes and point mutations were identified. To further understand the differences in resistance correlation between different sources, a presence/absence matrix was drawn to show the distribution of resistance genes and point mutations (Fig. 2). Of all the resistance genes and point mutations detected, those encoding fluoroquinolones were the most common, including genes such as acrB, emrA, emrB, H-NS, and rsmA. This was followed by genes encoding tetracyclines, such as emrK, emrY, evgA, and evgS, which were detected in the genomes of all strains.
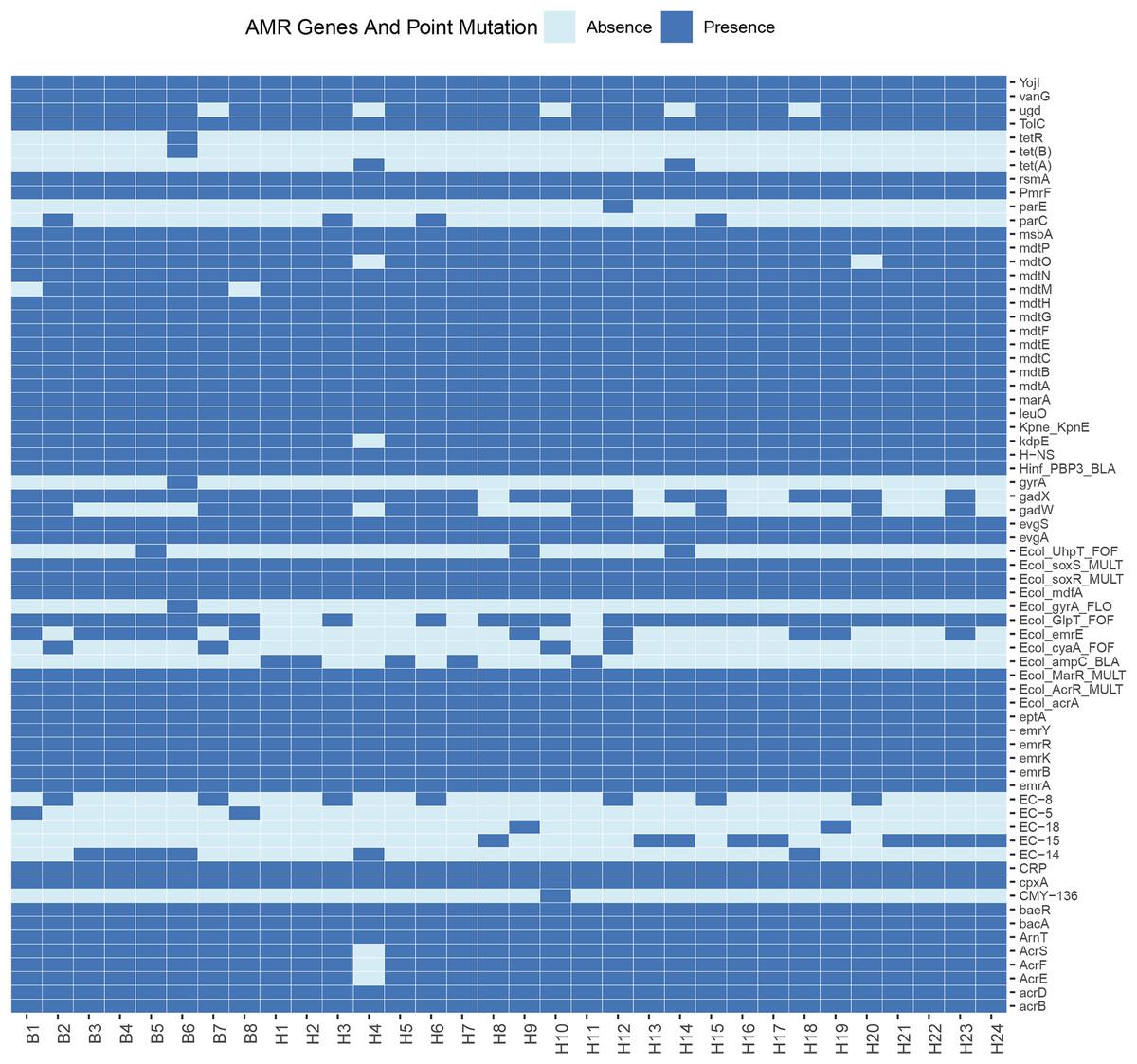
Figure 2: Profiles of AMR genes and point mutation for each strain in this study.
{kind=link}
A total of 111 virulence factors were detected in 32 E. coli isolates. To further understand the differences in virulence correlation between different sources, a presence/absence matrix was drawn to show the distribution of virulence genes (Fig. 3). The presence/absence matrix indicates that most strains have similar virulence genes. These virulence genes contribute to invasion, adherence, immune evasion, efflux pump, toxin, motility, stress adaption, and other virulence-related functions of E. coli.
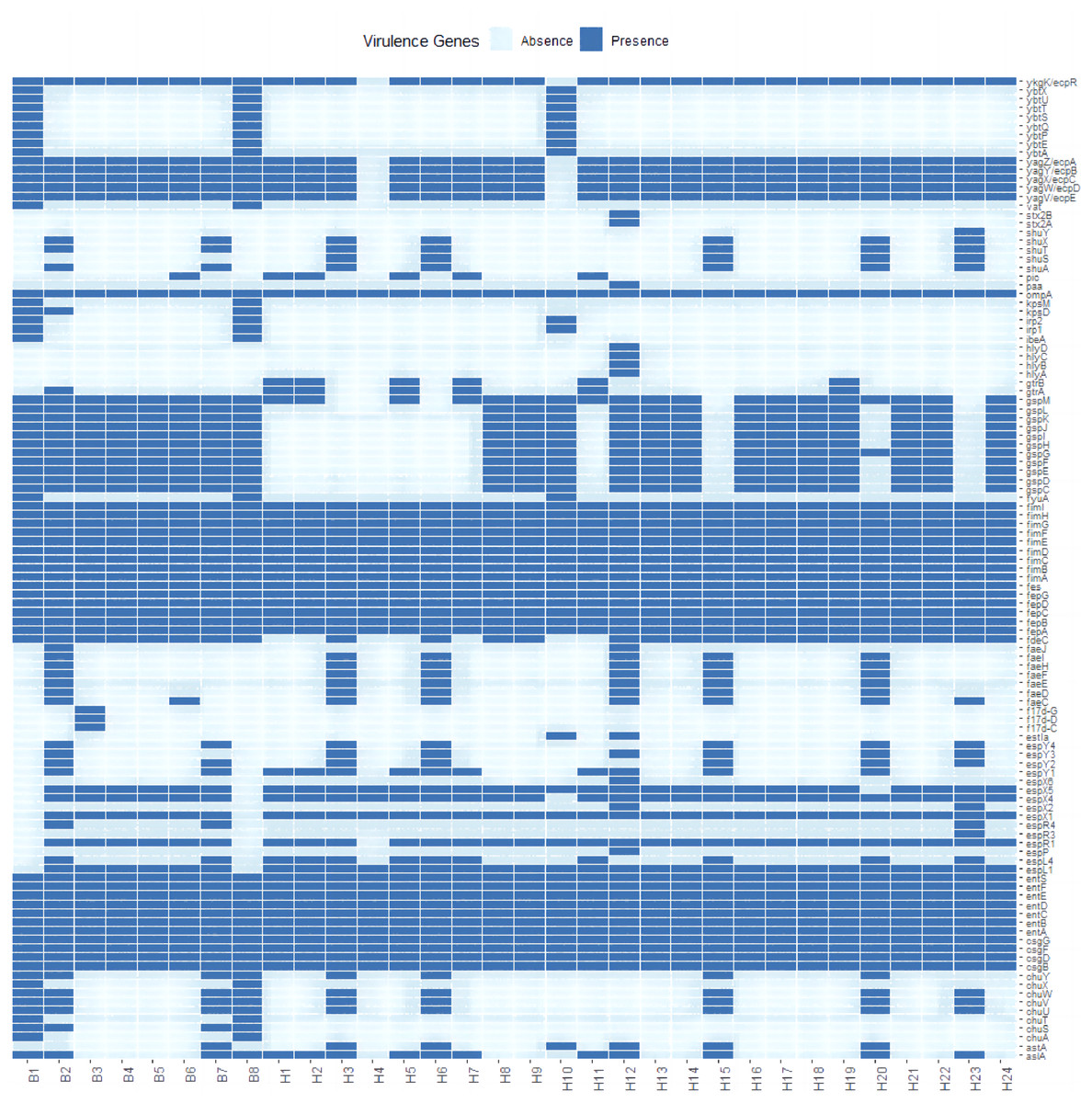
Figure 3: Profiles of virulence genes for each strain in this study.
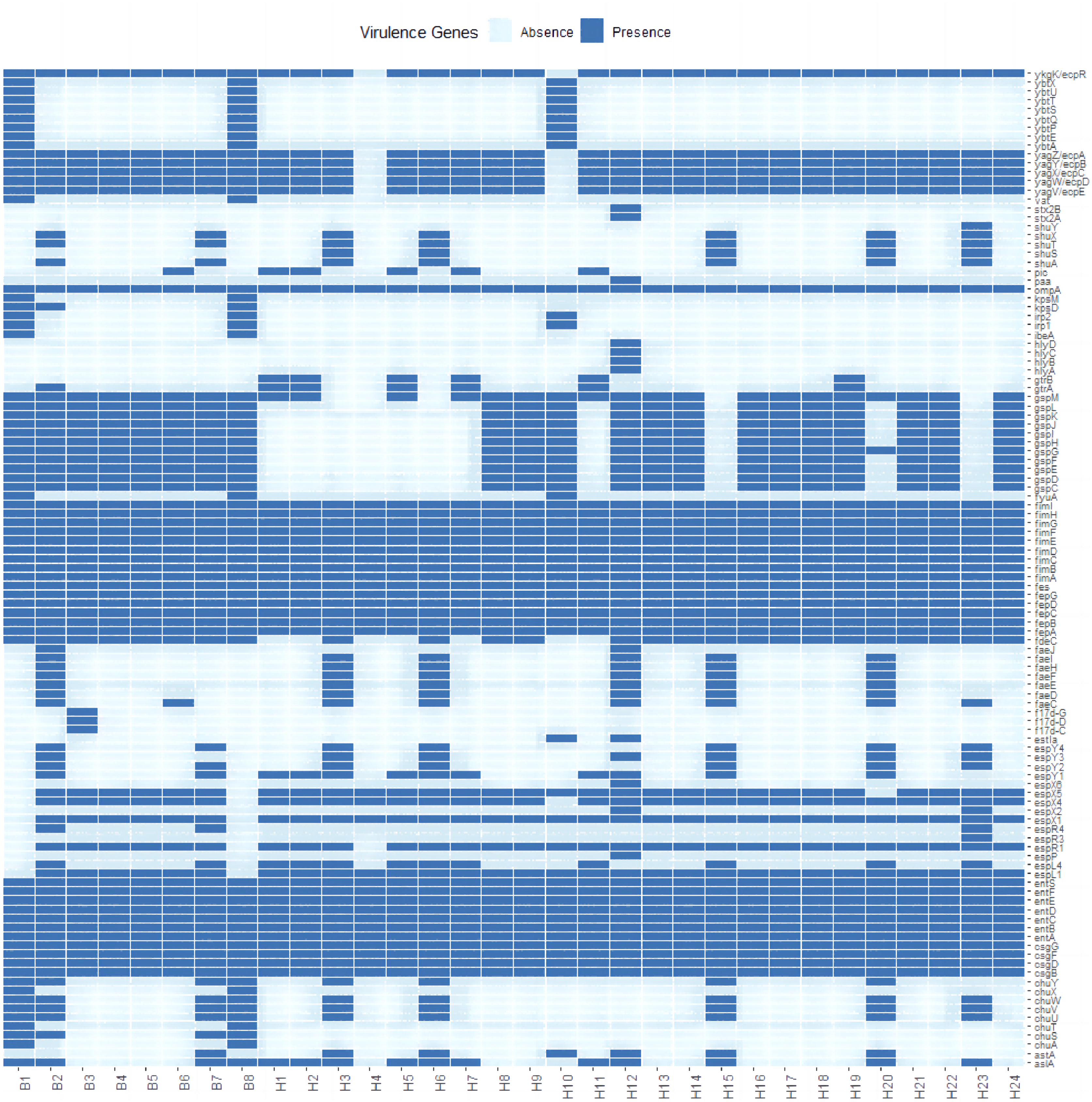{kind=link}
Phylogenetic analysis
The evolutionary relationships of the core genomic SNPs of 32 E. coli strains from wild tiger leopard feces are shown in Fig. 1. The maximum SNP distance between each strain was 59,699 SNPs, and the minimum distance was four SNPs. According to the branch lengths, the E. coli isolates with serotype O174:H23 (H16, H17, H13, H22, H24, H21, H8) had closer evolutionary distances to each other than to the other branches, with the number of SNPs differing from 4 to 19 (Table S4). As expected, closely grouped in the generated tree were E. coli isolates with the same sequence type and close or identical serotypes (Fig. 1). The phylogenetic tree based on core genomic SNPs was used to further investigate the transmission of strains ST939 and ST10 among different animals. From the evolutionary tree, it can be seen that E. coli with sequence types ST939 and ST10 are also present in other animals and clustered in a team with E. coli with sequence types ST939 and ST10 carried by wild Amur tigers and North China leopards (Fig. 4).
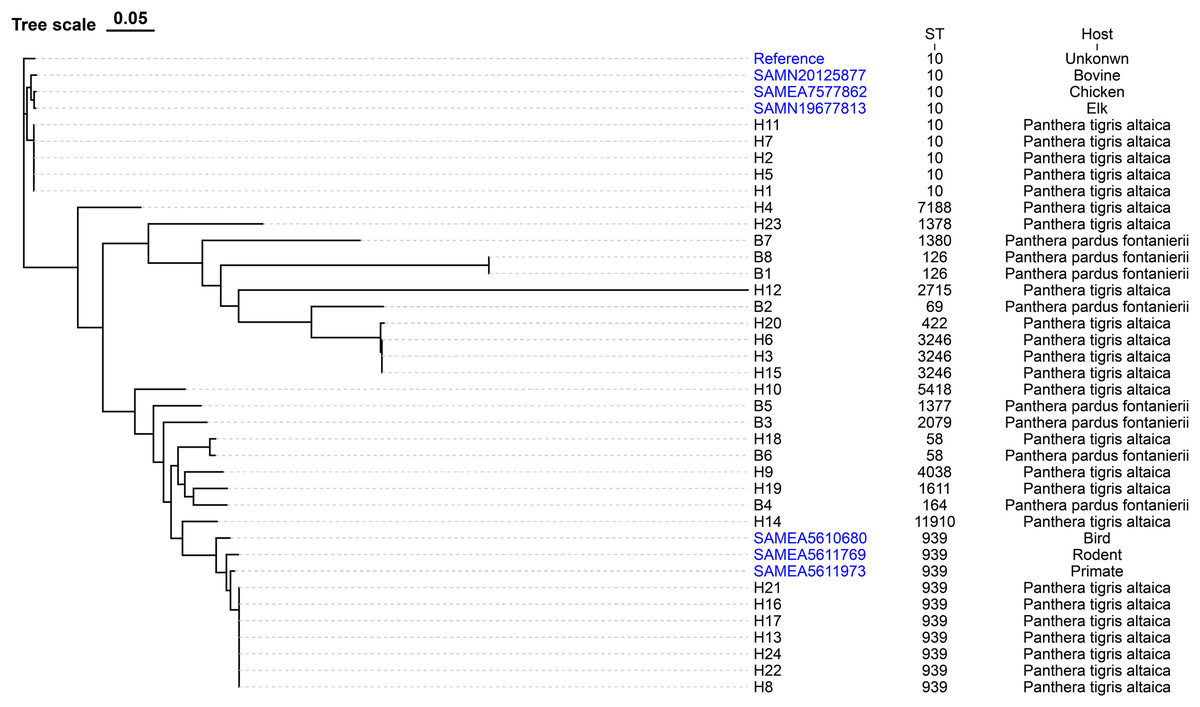
Figure 4: Phylogenetic tree of E. coli strains based on core genome SNPs.
The data used to construct the evolutionary tree include sequenced samples from 32 E. coli strains carried by wild Amur tigers and North China leopards, as well as some whole-genome data of E. coli with sequence types ST939 and ST10 downloaded from NCBI, with the reference sequence of E. coli MG1655. Blue color indicates that the sample was downloaded from NCBI.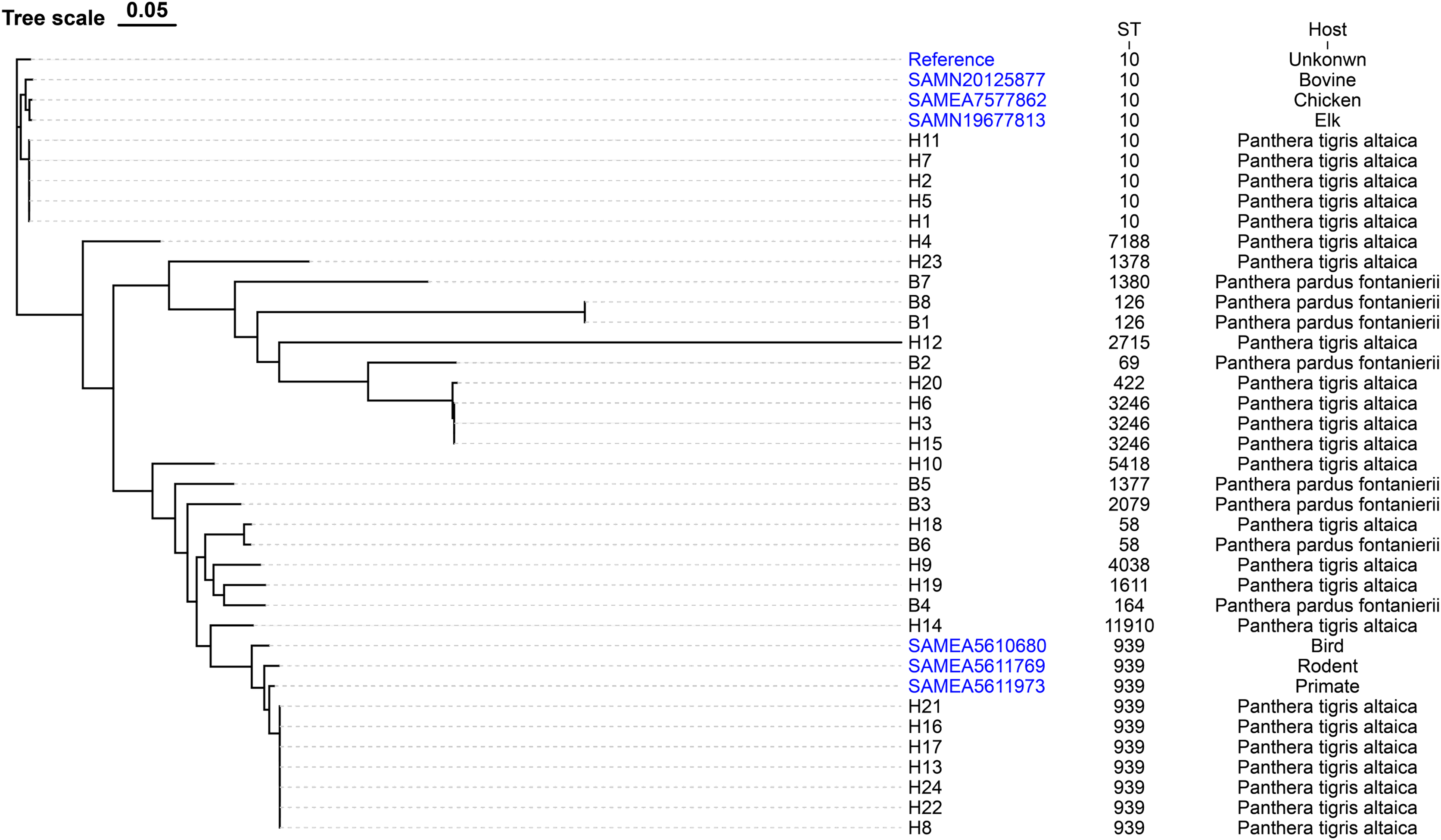{kind=link}
Discussion
E. coli is widely present in nature, water and food, as well as the respiratory tract and intestinal tract of animals in the normal flora, as the intestinal resident bacteria, in the deterioration of conditions and other pathogenic infections are easy to follow the disease. Some resistance studies have found widespread resistance in E. coli from captive tiger sources, and the susceptibility of different pathogenic E. coli strains to antibiotics varies widely, including resistance to ampicillin, gentamicin, and chloramphenicol (Qiu et al., 2016). Here we performed whole-genome sequencing of 32 strains isolated from wild Amur tiger and North China leopard feces. Among these isolates, we detected 68 resistance genes and point mutations. Of all the resistance genes and point mutations detected, genes encoding fluoroquinolones were the most prevalent, including the genes acrB, emrA, emrB, H-NS, and rsmA. This was followed by genes encoding tetracyclines, such as emrK, emrY, evgA, and evgS, which were detected in the genomes of all strains. Notably, genes from the exocytosis pump family accounted for the largest proportion of the detected resistance genes. This result suggests that the efflux pumps may be associated with antibiotic resistance in the investigated strains. The results of the antimicrobial susceptibility testing showed that among the 32 E. coli isolates, there were significant differences in the resistance rates to different drugs. Specifically, the resistance rate to tetracycline was relatively high at 43.8%, but the resistance rates to imipenem, doxycycline, norfloxacin, and ciprofloxacin were relatively low, none of which exceeded 10%. This result suggests that drug-resistant E. coli has appeared in the wild populations of the Amur Tigers and the North China Leopards.
In recent years, as more and more virulence factors are understood in E. coli, the pathogenic processes of some of these virulence factors have increasingly come into focus. The pathogenicity of E. coli is the result of the synergistic effect of multiple virulence factors, and some studies have shown that E. coli of tiger origin has a variety of virulence factors, including Stx2f, fimC, cnf-1, fyuA, iroc, irp2, etc., which may be related to the severity of the infection and the course of the disease may also be an important cause of the disease (Zhang, 2022). By comparison with the virulence factor database, 111 virulence genes were identified in E. coli isolates. The virulence genes with the highest detection rates included the genes fimA, fimB, fimC, fimD, fimE, fimF and fimH, which were detected in the genomes of all strains. We also detected the stx2A and stx2B gene, which produces Shiga toxin, in an E. coli isolate. Clinical symptoms of Shiga toxin-producing E. coli infections can include hemorrhagic colitis, bloody or watery diarrhea, and potentially fatal hemolytic uremia (HUS), which can lead to sudden kidney failure. However, it is unclear whether these pathogenic E. coli cause disease in these endangered wild Amur tiger and North China leopard, or whether they contribute to the continued decline of populations. It is clear that further research into the microbial diversity of this Amur tiger and North China leopard should be undertaken as part of future investigations to aid their recovery efforts. In this study, whole genome sequencing data showed that 32 E. coli isolates were highly serotypically diverse, with a total of 18 different serotypes identified, including four isolates with no O serotypes identified, and those belonging to the same serotype showed extensive similarity and clustered together phylogenetically. The pathogenicity of E. coli is linked to the type of O-antigen serotype, with different pathogenic E. coli having different dominant O-antigen serotypes. In our study, we identified the dominant serotype O1 of E. coli associated with urinary tract infections and the dominant serotype O125ab of enteropathogenic E. coli. with serotype O125, first identified in 1952 during a diarrheal outbreak in London (Taylor & Charter, 1952) has since been isolated from diarrheal patients worldwide (Croxen et al., 2013) and has been recognized as an important pathogenic EPEC serotype by the World Health Organization. However, it is unclear whether these pathogenic serotypes of E. coli cause disease in endangered wild Amur tigers and North China leopards, or whether they lead to a sustained population decline. Clearly, research on E. coli pathogenicity carried by Amur tigers and North China leopards should be further deepened as part of future investigations.
Conclusions
In this study, E. coli from wild Amur tigers and North China leopards were analyzed by whole genome sequencing, and we detected multiple drug resistance genes in E. coli isolates. We also identified several virulence factors with high pathogenicity as well as serotypes, including the Stx2A and Stx2B gene that produces Shiga toxin, as well as the dominant serotype O1 of E. coli associated with urinary tract infections and the dominant serotype O125ab of intestinal pathogenic E. coli. At the same time, through antimicrobial susceptibility testing, we found that some of these E. coli isolates were resistant to antibiotics such as tetracycline and imipenem. These would be a potential threat to wild Amur tigers and North China leopards. Our findings, therefore, highlight the importance of surveillance programs, particularly given the potential impact on endangered wildlife and human public health.