
An early geikiid dicynodont from the Tropidostoma Assemblage Zone (late Permian) of South Africa
- Published
- Accepted
- Received
- Academic Editor
- Andrew Farke
- Subject Areas
- Evolutionary Studies, Paleontology, Taxonomy, Zoology
- Keywords
- Synapsida, Therapsida, Dicynodontia, Permian, Karoo Basin, South Africa, Ghost lineage, Phylogeny
- Copyright
- © 2017 Kammerer et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2017. An early geikiid dicynodont from the Tropidostoma Assemblage Zone (late Permian) of South Africa. PeerJ 5:e2913 https://doi.org/10.7717/peerj.2913
Abstract
Based on specimens previously identified as Tropidostoma, a new taxon of dicynodont (Bulbasaurus phylloxyron gen. et sp. nov.) from the Karoo Basin of South Africa is described. Bulbasaurus is a medium-sized dicynodont (maximum dorsal skull length 16.0 cm) restricted to the Tropidostoma Assemblage Zone (early Lopingian) of the Beaufort Group. Bulbasaurus can be distinguished from Tropidostoma by an array of characters including the presence of a tall, sharp premaxillary ridge, large, rugose, nearly-confluent nasal bosses, a nasofrontal ridge, massive tusks, robust pterygoids, prominently twisted subtemporal bar, and absence of a distinct postfrontal. Inclusion of Bulbasaurus in a phylogenetic analysis of anomodont therapsids recovers it as a member of Geikiidae, a clade of otherwise later Permian dicynodonts such as Aulacephalodon and Pelanomodon. Bulbasaurus exhibits many of the characters typical of adult Aulacephalodon, but at substantially smaller skull size (these characters are absent in comparably-sized Aulacephalodon juveniles), suggesting that the evolution of typical geikiid morphology preceded gigantism in the clade. Bulbasaurus is the earliest known geikiid and the only member of the group known from the Tropidostoma Assemblage Zone; discovery of this taxon shortens a perplexing ghost lineage and indicates that abundant clades from the later Permian of South Africa (e.g., Geikiidae, Dicynodontoidea) may have originated as rare components of earlier Karoo assemblage zones.
Introduction
Several major turnovers in tetrapod faunal composition occurred in the Karoo Basin of South Africa during the span of time recorded in the Permo-Triassic Beaufort Group (Rubidge, 1995). The best-known and most intensely-studied of these turnovers corresponds to the end-Permian mass extinction, which wiped out the majority of species and several major clades of tetrapods (Smith & Ward, 2001; Retallack, Smith & Ward, 2003; Ward et al., 2005; Botha-Brink & Angielczyk, 2010; Irmis & Whiteside, 2012; Fröbisch, 2013; Smith & Botha-Brink, 2014). Recently, however, increased research attention has also been given to the turnover between middle (Guadalupian) and late (Lopingian) Permian Karoo faunas. Studies of this transition have primarily focused on the role of extinction in driving faunal change, and the possible relationship between Karoo tetrapod turnover and global marine turnover as part of a hypothesized mid-Permian mass extinction (Rubidge et al., 2013; Rubidge et al., 2016; Day et al., 2015a; Day et al., 2015b; Réy et al., 2016). However, the primary victims of mid-Permian extinction among Karoo tetrapods were the dinocephalian therapsids, a group that, although ecologically diverse and species-rich (Boonstra, 1969; Kammerer, 2011), represent a relatively small component of middle Permian faunas in terms of specimen abundance (Smith, Rubidge & van der Walt, 2012). The most abundant middle Permian Karoo therapsids were, just as in the late Permian and earliest Triassic, the anomodonts, and specifically their subclade Dicynodontia. Dicynodonts suffered minimal extinction-related turnover between the middle and late Permian. Indeed, the most abundant middle Permian dicynodont (the pylaecephalid Diictodon feliceps) only becomes more abundant in late Permian strata (Smith, Rubidge & van der Walt, 2012) (before finally trailing off and going extinct in the terminal Permian Daptocephalus Assemblage Zone; Viglietti et al., 2016). The next-most-abundant small-bodied Permian dicynodonts (Pristerodon and Emydops) also first appear in the middle Permian (Angielczyk, Fröbisch & Smith, 2005; Kammerer, Angielczyk & Fröbisch, 2011) and survive the mid-Permian extinction seemingly unscathed. Among dicynodont taxa whose ranges do not extend into the late Permian, most are components of the earliest Karoo faunal assemblages, none of whose members even make it to the end of the middle Permian (e.g., Eodicynodon, Lanthanostegus, Colobodectes) (Modesto, Rubidge & Welman, 2002; Angielczyk & Rubidge, 2009). Furthermore, the few dicynodont taxa that are apparent victims of the mid-Permian extinction (Brachyprosopus broomi and Robertia broomiana) have close relatives (at the ‘family level’) that thrive in the late Permian (Angielczyk & Rubidge, 2013; Cox & Angielczyk, 2014; Angielczyk et al., 2016).
Even though dicynodonts show robust survivorship between the middle and late Permian, there are nonetheless major differences between middle and late Permian Karoo dicynodont faunas. However, these differences are not driven by extinction within the clade, but rather by origination, and specifically the appearance of large-bodied taxa. All middle Permian dicynodonts are relatively small (skull length <20 cm) animals; the niche of large herbivore in the middle Permian Karoo was restricted to dinocephalians and pareiasaurs (Boonstra, 1969). In the late Permian, however, dicynodonts are overwhelmingly the most abundant large herbivores (Kammerer, Angielczyk & Fröbisch, 2011; Smith, Rubidge & van der Walt, 2012). Although this may have been driven in part by the opening up of ecospace resulting from the extinction of herbivorous dinocephalians, there is a substantial lag between dinocephalian extinction and large dicynodont evolution. Dinocephalians go extinct at the end of the Tapinocephalus Assemblage Zone (AZ) (Day et al., 2015a). Only one large-bodied dicynodont (the aberrant Endothiodon bathystoma; Cox & Angielczyk, 2014) appears immediately in the wake of dinocephalian extinction, in the overlying Pristerognathus AZ, and few large-bodied dicynodonts are present in the subsequent Tropidostoma AZ. The majority of large-bodied dicynodont taxa do not appear until the later Cistecephalus and Daptocephalus AZs, roughly five million years afterwards (Smith, Rubidge & van der Walt, 2012; Rubidge et al., 2013; Viglietti et al., 2016).
With the notable exception of the aforementioned Endothiodon, almost all large (>30 cm skull length) dicynodont herbivores belong to the clade Bidentalia. Bidentalians are characterized by a reduction in dentition relative to earlier dicynodonts (in almost all taxa only the maxillary tusks are present, if teeth are present at all) and include a number of abundant and well-known Karoo taxa such as Dicynodon, Daptocephalus, Oudenodon, and Aulacephalodon (Kammerer & Angielczyk, 2009). In South Africa, this clade first appears in the Tropidostoma AZ, in the form of the eponymous Tropidostoma dubium, a medium-sized oudenodontid (adult skull length: ∼20 cm; Botha & Angielczyk, 2007). The Tropidostoma AZ is a crucial turning-point for Karoo therapsid faunas in general: cynodonts, which would go on to become the most successful Mesozoic therapsids, first appear in this assemblage zone (Botha, Abdala & Smith, 2007; Kammerer, 2016), and eutherocephalians and gorgonopsians, the dominant predators of the later Permian, show a marked increase in abundance (Smith, Rubidge & van der Walt, 2012; Kammerer et al., 2015). The primary bidentalian component of the Tropidostoma AZ is Tropidostoma itself, although rare records of Oudenodon and Rhachiocephalus also appear in the upper part of this assemblage zone (Botha & Angielczyk, 2007; Smith, Rubidge & van der Walt, 2012). Perplexing in their absence, however, are any members of the clades Dicynodontoidea or Geikiidae. Dicynodontoids represent one of the two major bidentalian subclades (the other being Cryptodontia, which includes the aforementioned Tropidostoma, Oudenodon, Rhachiocephalus, and allied taxa) and are very conspicuous members of later assemblage zones (including the nominal genera for the Daptocephalus and Lystrosaurus AZs). Geikiids are a cryptodont group, and include the abundant and highly autapomorphic large-bodied taxon Aulacephalodon bainii, which first appears in the Cistecephalus AZ (Tollman, Grine & Hahn, 1980; Kammerer, Angielczyk & Fröbisch, 2011; Kammerer, Angielczyk & Fröbisch, 2015a).
Based on the presence of their sister-clades in the Tropidostoma AZ, dicynodontoids and geikiids must have diverged by this time. There are three main explanations for the absence of these taxa in the Tropidostoma AZ of South Africa. One is that these taxa were present but rare components of the fauna, and sampling effort has not been sufficient to recover their remains. However, the Tropidostoma AZ is a very well-sampled assemblage zone (Smith, Rubidge & van der Walt, 2012), and has already yielded specimens of even extremely rare therapsid taxa such as burnetiamorphs (Sidor, Hopson & Keyser, 2004; Sidor & Smith, 2007). A second explanation is that the missing taxa did not originate in South Africa, and only dispersed to the Karoo Basin later in their evolution. This is an important possibility to consider, given how little we know about late Permian tetrapod faunas globally, and the fact that bidentalians were already also present in Russia in probable Tropidostoma AZ-equivalent strata (Kurkin, 2000; Angielczyk & Kurkin, 2003). There is a third possibility, however—that these taxa were present in the Tropidostoma AZ and their fossils have been collected, but not recognized as such. South African dicynodont alpha taxonomy has only recently been stabilized after roughly a century in the wilderness (see Kammerer, Angielczyk & Fröbisch (2011) for a review). In the absence of meaningful species diagnoses in the wake of Broomian hyper-splitting in the early 20th century, researchers working on Karoo biostratigraphy tended to refer newly-collected dicynodont specimens to a few, broadly-understood dicynodont genera (Kitching, 1977; Cluver & Hotton, 1981). Now that the majority of nominal taxa have been resolved into well-diagnosed morphospecies, these broad earlier specimen referrals can be reevaluated, and in some cases have proven to be over-lumped, creating heterogeneous arrays of unrelated taxa (Kammerer, Angielczyk & Fröbisch, 2011; Kammerer, Angielczyk & Fröbisch, 2015b). This is particularly problematic when dealing with basal members of highly apomorphic clades, i.e., taxa that do not yet show the full suite of synapomorphies traditionally used to characterize those clades. Specimens of such species may instead end up referred to coeval, generalized dicynodont taxa, obscuring their importance as early representatives of clades that would later become major components of their faunas (e.g., the earliest lystrosaurid Euptychognathus, originally referred to ‘Dicynodon’; Kammerer, Angielczyk & Fröbisch, 2011). Here, we present another such case, recognizing the first geikiid dicynodont from the Tropidostoma AZ on the basis of specimens previously identified as Tropidostoma itself.
Materials and Methods
All specimens mentioned in the text were examined personally by the lead author. For information on the methodology of the phylogenetic analysis, refer to the Discussion. The electronic version of this article in Portable Document Format (PDF) will represent a published work according to the International Commission on Zoological Nomenclature (ICZN), and hence the new names contained in the electronic version are effectively published under that Code from the electronic edition alone. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix http://zoobank.org/. The LSID for this publication is: urn:lsid:zoobank.org:pub:EB1EE1F8-C512-4272-BFA3-C8938E0806E5. The online version of this work is archived and available from the following digital repositories: PeerJ, PubMed Central and CLOCKSS.
The holotype specimen of the new taxon (SAM-PK-K11235) was collected by one of the authors (RMHS) in Tropidostoma AZ exposures of the locality Vredelus (Fraserburg District, Western Cape Province). The Tropidostoma AZ strata in the southern Karoo Basin mainly coincide with the argillaceous Hoedemaker Member (Teekloof Formation), which represents the distal portion of a large distributary fluvial system. Three dimensional outcrops of exhumed paleomeanderbelts (the Reiersvlei Sandstone of Smith (1987)) in the vicinity of the type locality of the new dicynodont show that the former rivers had channel widths of up to 350 m, and point bar diameters of ∼3 km.

Figure 1: SAM-PK-K11235, holotype of Bulbasaurus phylloxyron gen. et sp. nov., in situ at the type locality of Driekoppe, Vredelus, Fraserburg, Western Cape Province, South Africa.
(A) Specimen in left semi-lateral view; (B) specimen in dorsal view.{kind=link}
SAM-PK-K11235 was preserved dorsal-side-up in a 1.5 m thick bed of structureless grey siltstone (Fig. 1) with scattered small micrite nodules immediately overlying a 1 m-thick fine-grained ripple cross-laminated sandstone with mudrock rip-up clasts. The massive siltstone is interpreted as having been deposited on the proximal floodplain alongside a large northeasterly-flowing meandering river on one of the low angled fluvial fans that emanated from the Gondwanide mountains. The linear channel-fill sandstone beneath represents a minor floodplain distributary.
The holotype skull was found in situ with no skeletal elements in association and was not perimineralized with calcareous nodular material. This ‘head-only’ taphonomic style is typical of therapsid occurrences in proximal floodplain deposits of the Hoedemaker Member (Smith, 1993) and is attributed to extended periods of non-deposition, allowing for sub-aerial disarticulation of skeletons, then short transportation and rapid burial by silt-laden overbank floods.
SAM-PK-K11235 was mechanically prepared by Mr. Sibusiso Mtungata in the Karoo palaeontology laboratory, Iziko Museums of South Africa (Cape Town). All other specimens referred to the new dicynodont taxon were also mechanically prepared, either at Iziko or the Council for Geoscience (Pretoria).
Systematic Paleontology
| Synapsida Osborn, 1903 |
| Therapsida Broom, 1905 |
| Anomodontia Owen, 1860 |
| Dicynodontia Owen, 1860 |
| Cryptodontia Owen, 1860 |
| Geikiidae Nopcsa, 1923 |
Type genus: Geikia Newton, 1893.
Included genera: Aulacephalodon Seeley, 1898; Bulbasaurus gen. nov.; Pelanomodon Broom, 1938.
Bulbasaurus gen. nov.
LSID: urn:lsid:zoobank.org:act:F21D10AF-277A-44D4-B374-BE89AF2B7541
Type species: Bulbasaurus phylloxyron sp. nov.
Etymology: From the Latin bulbus (a bulb), referring to the shape of the nasal boss, and the latinized Greek saurus (a lizard), a common suffix for fossil amniotes of ‘reptilian’ grade.
Diagnosis: As for the type and only species.
Bulbasaurus phylloxyron sp. nov.
LSID: urn:lsid:zoobank.org:act:C80D31CA-86B1-43C4-9EA9-CED4A5852262
Holotype: SAM-PK-K11235, a partial skull missing the left subtemporal bar and both postorbital bars (Figs. 1–6).
Type locality and horizon: Driekoppe, Vredelus, Fraserburg, Western Cape Province, South Africa. Hoedemaker Member of the Teekloof Formation, ∼50 m above top of Reiersvlei Sandstone at a height of 1,445 m, Tropidostoma Assemblage Zone (Lopingian).

Figure 2: SAM-PK-K11235, holotype of Bulbasaurus phylloxyron gen. et sp. nov., in (A) anterior, (B) right semi-lateral, and (C) left lateral views.
Scale bar equals 5 cm.{kind=link}
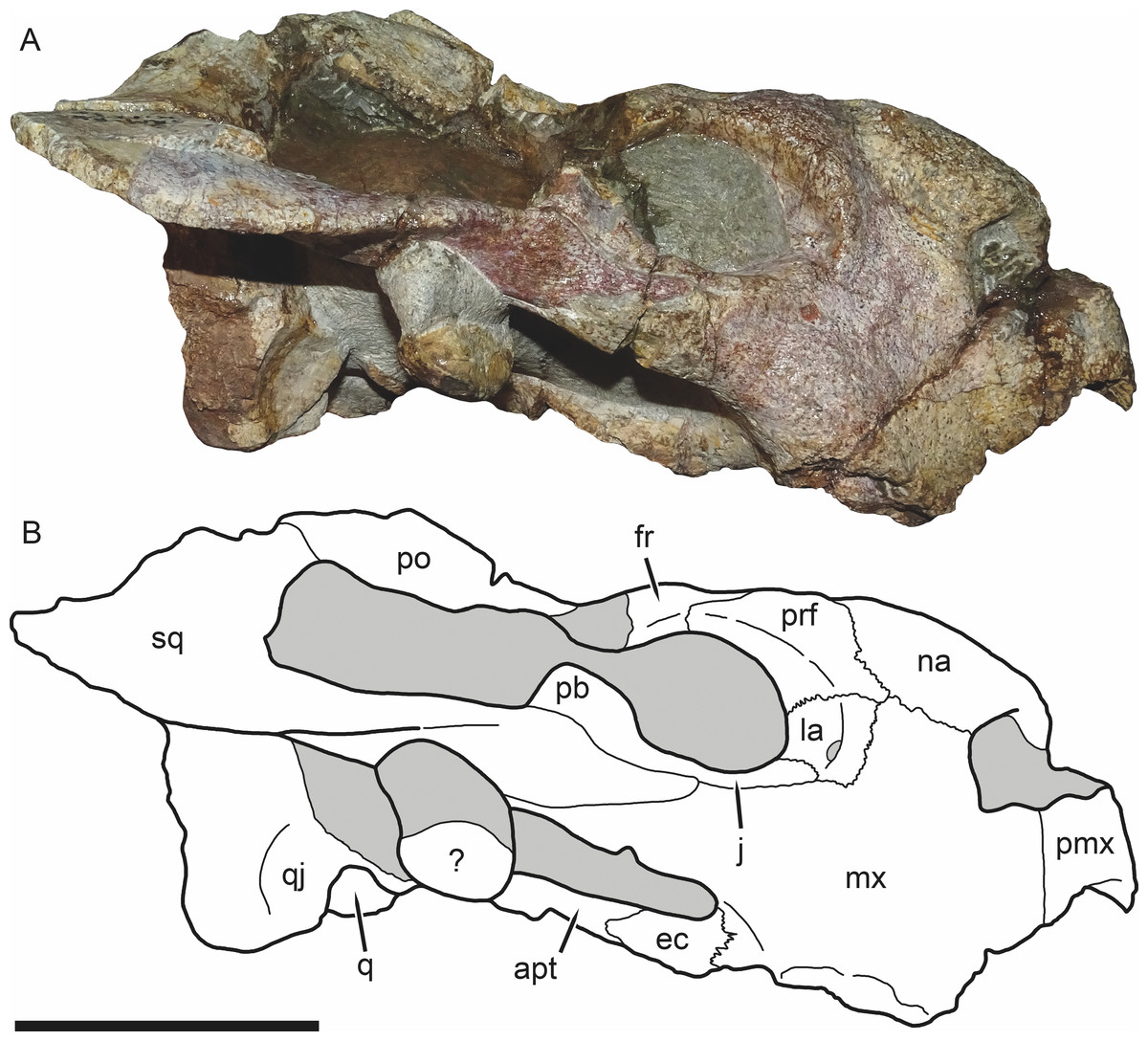
Figure 3: SAM-PK-K11235, holotype of Bulbasaurus phylloxyron gen. et sp. nov., in right lateral view.
(A) photograph and (B) interpretive drawing. Abbreviations: ?, unknown bone; apt, anterior pterygoid ramus; ec, ectopterygoid; fr, frontal; la, lacrimal; mx, maxilla; na, nasal; pb, base of postorbital bar; pmx, premaxilla; po, postorbital; prf, prefrontal; q, quadrate; qj, quadratojugal; sq, squamosal. Gray indicates matrix. Scale bar equals 5 cm.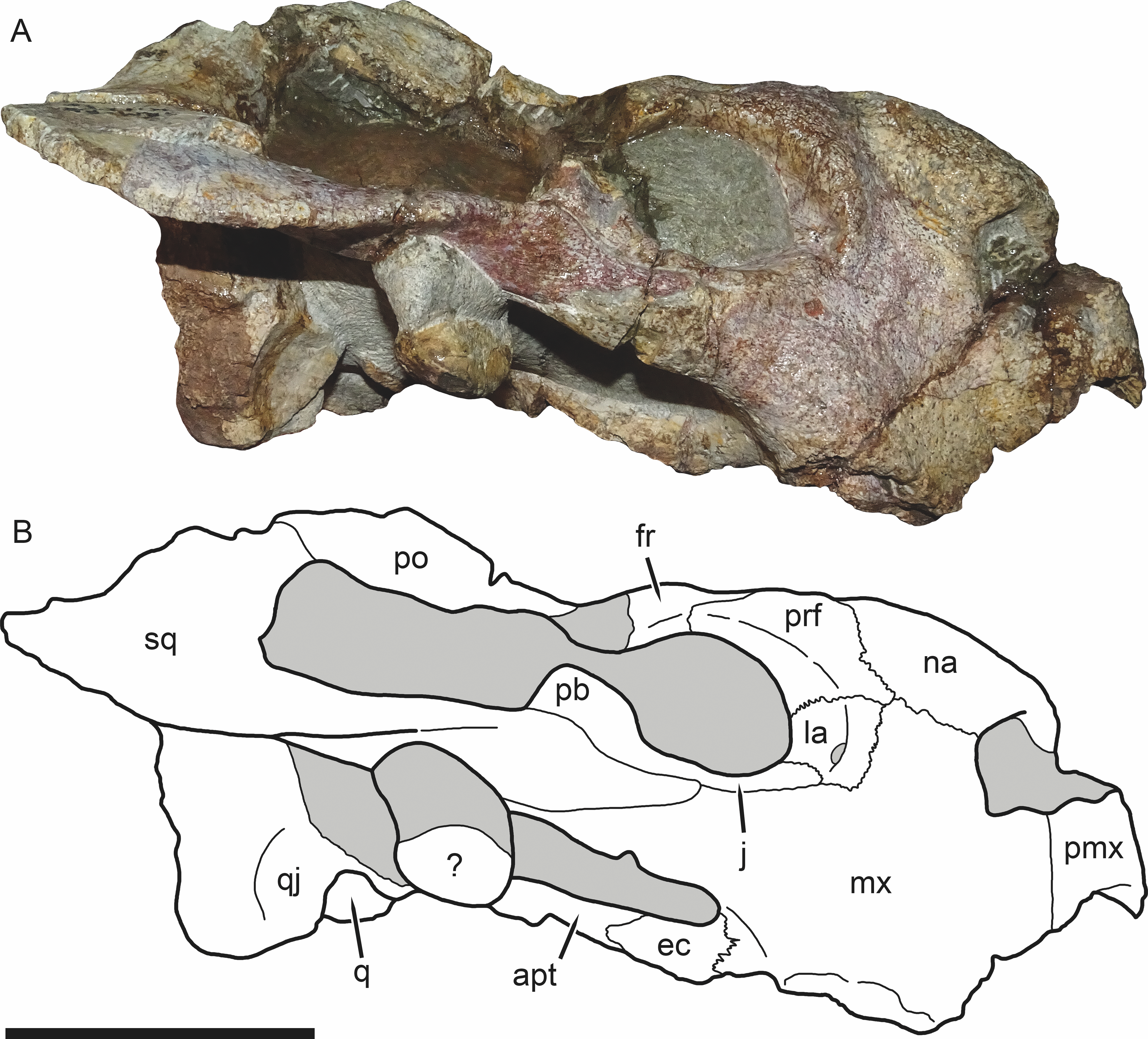{kind=link}
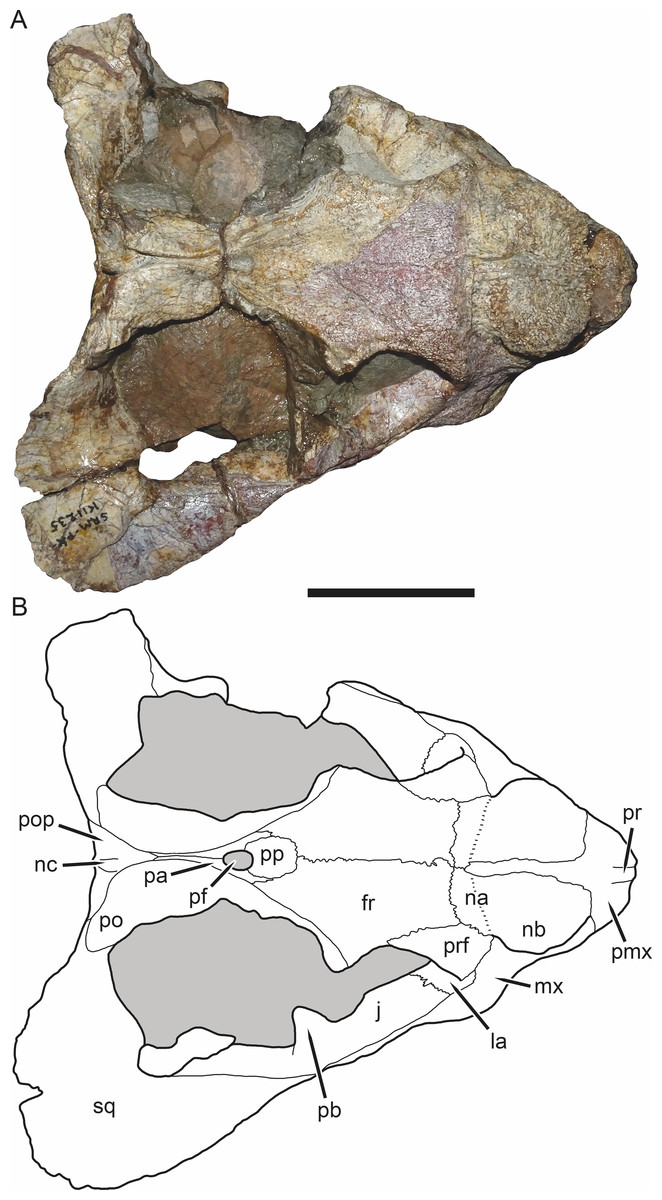
Figure 4: SAM-PK-K11235, holotype of Bulbasaurus phylloxyron gen. et sp. nov., in dorsal view.
(A) photograph and (B) interpretive drawing. Abbreviations: fr, frontal; la, lacrimal; j, jugal; na, nasal; nb, nasal boss; nc, nuchal crest; pa, parietal; pb, base of postorbital bar; pf, pineal foramen; pmx, premaxilla; po, postorbital; pop, postparietal; pp, preparietal; pr, premaxillary ridge; prf, prefrontal; sq, squamosal. Gray indicates matrix. Scale bar equals 5 cm.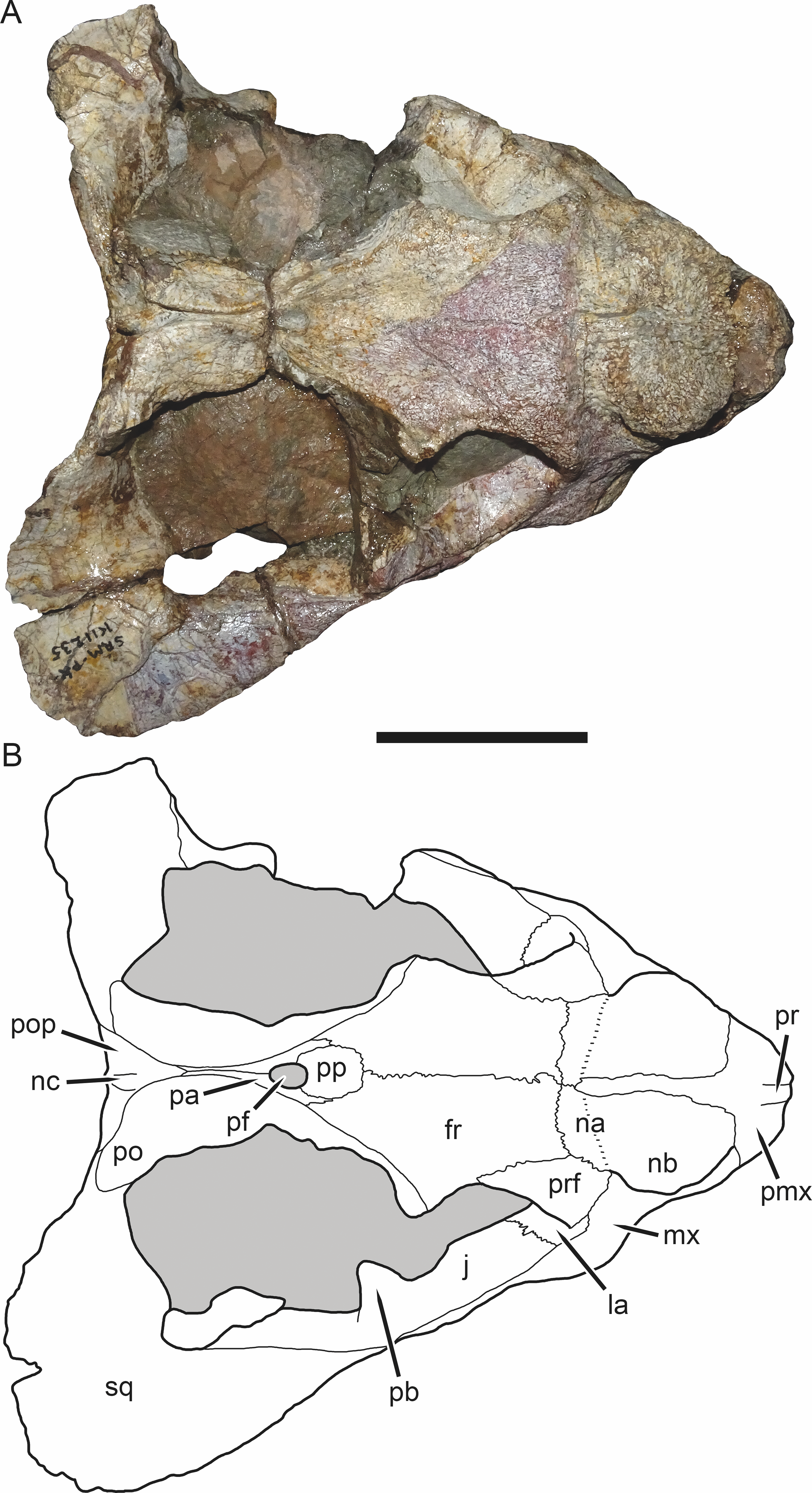{kind=link}
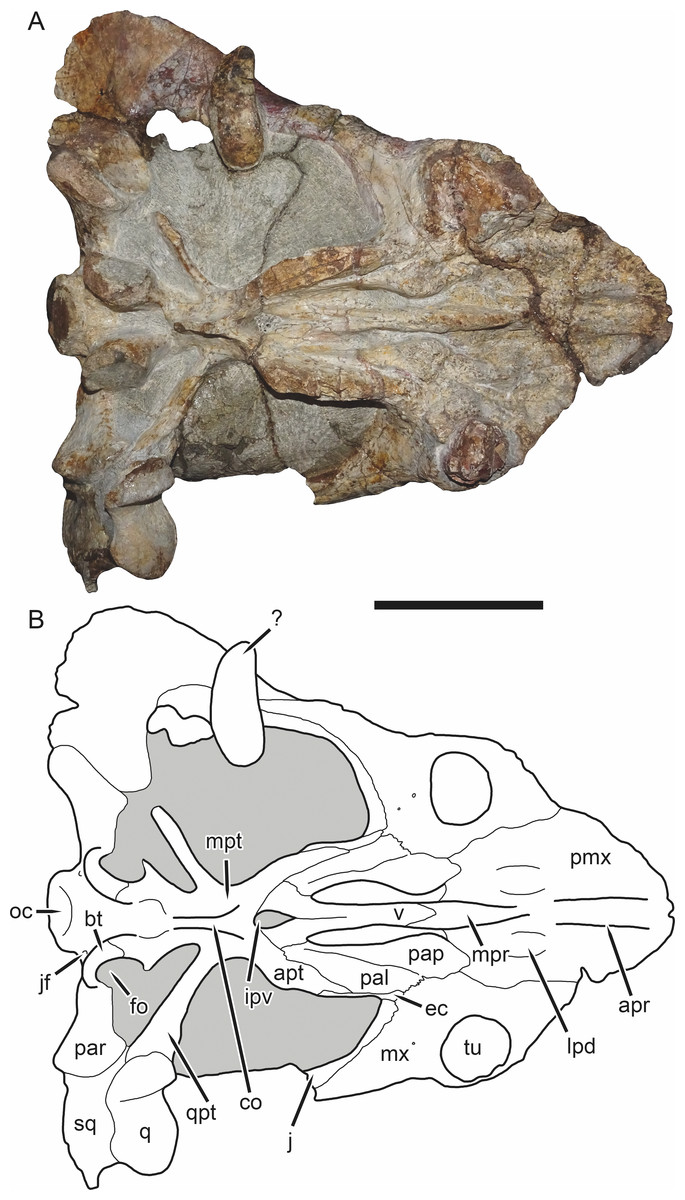
Figure 5: SAM-PK-K11235, holotype of Bulbasaurus phylloxyron gen. et sp. nov., in ventral view.
(A) photograph and (B) interpretive drawing. Abbreviations: ?, unknown bone; apr, anterior palatal ridge; apt, anterior pterygoid ramus; bt, basal tuber; co, crista oesophagea; ec, ectopterygoid; fo, fenestra ovalis; ipv, interpterygoid vacuity; j, jugal; jf, jugular foramen; lpd, lateral premaxillary depression; mpr, median palatal ridge; mpt, median pterygoid plate; mx, maxilla; oc, occipital condyle; pal, palatine; pap, palatine pad; par, paroccipital process; pmx, premaxilla; q, quadrate; qpt, quadrate ramus of pterygoid; sq, squamosal; tu, tusk; v, vomer. Gray indicates matrix. Scale bar equals 5 cm.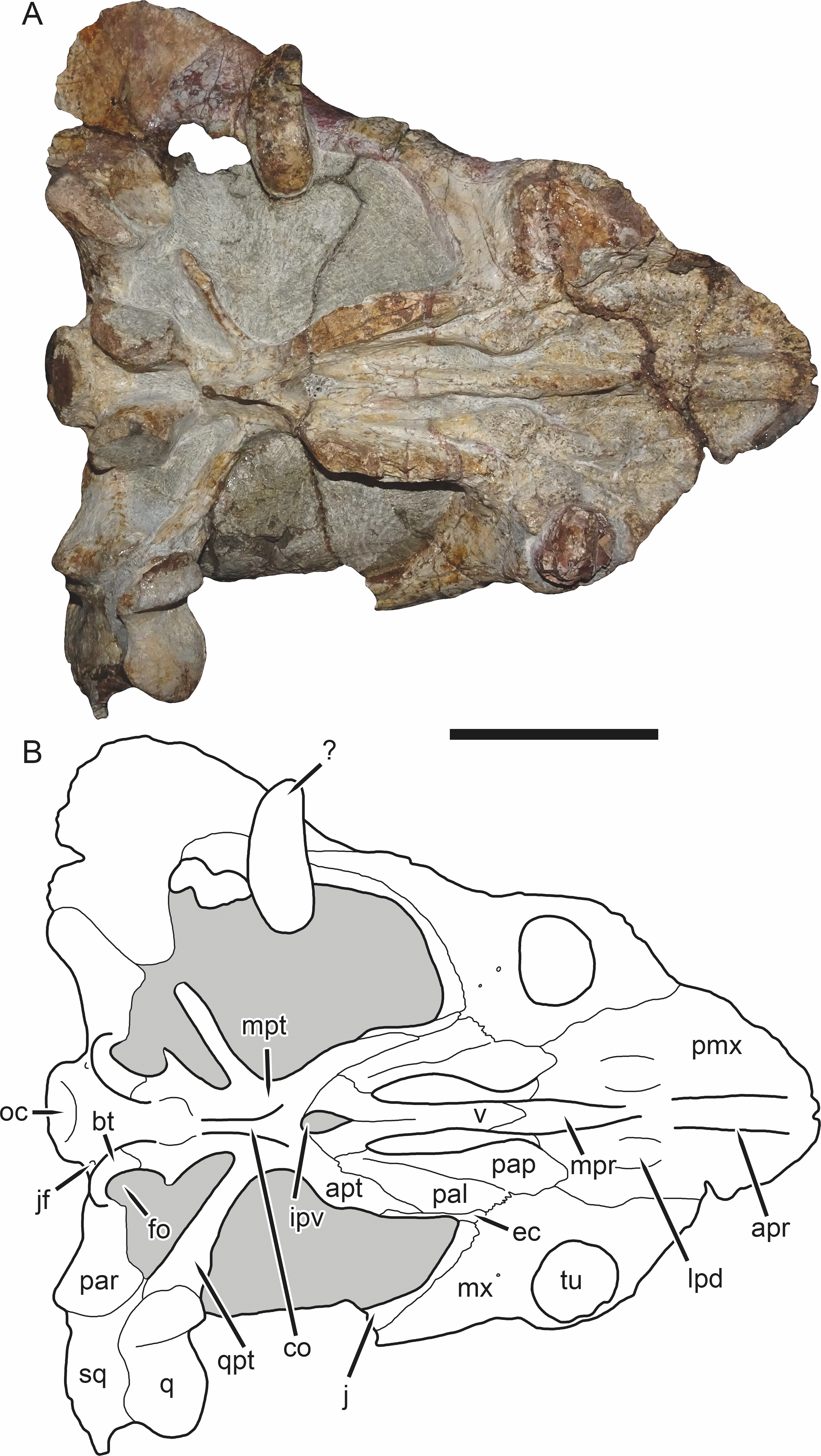{kind=link}
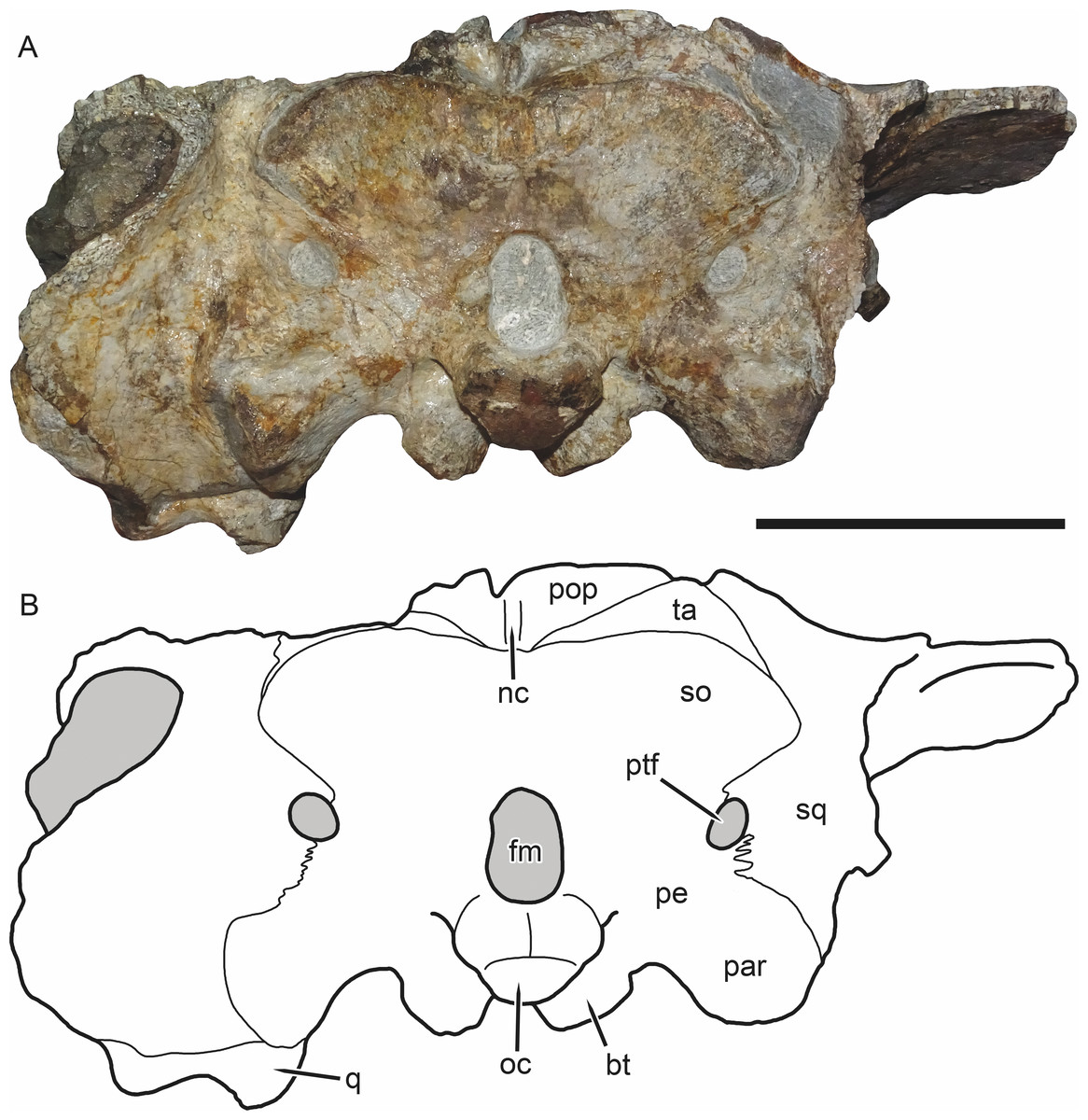
Figure 6: SAM-PK-K11235, holotype of Bulbasaurus phylloxyron gen. et sp. nov., in occipital view.
(A) photograph and (B) interpretive drawing. Abbreviations: bt, basal tuber; fm, foramen magnum; nc, nuchal crest; oc, occipital condyle; par, paroccipital process; pe, periotic; pop, postparietal; ptf, post-temporal fenestra; q, quadrate; so, supraoccipital (exact boundaries uncertain, fused with periotic); sq, squamosal; ta, tabular. Gray indicates matrix. Scale bar equals 5 cm.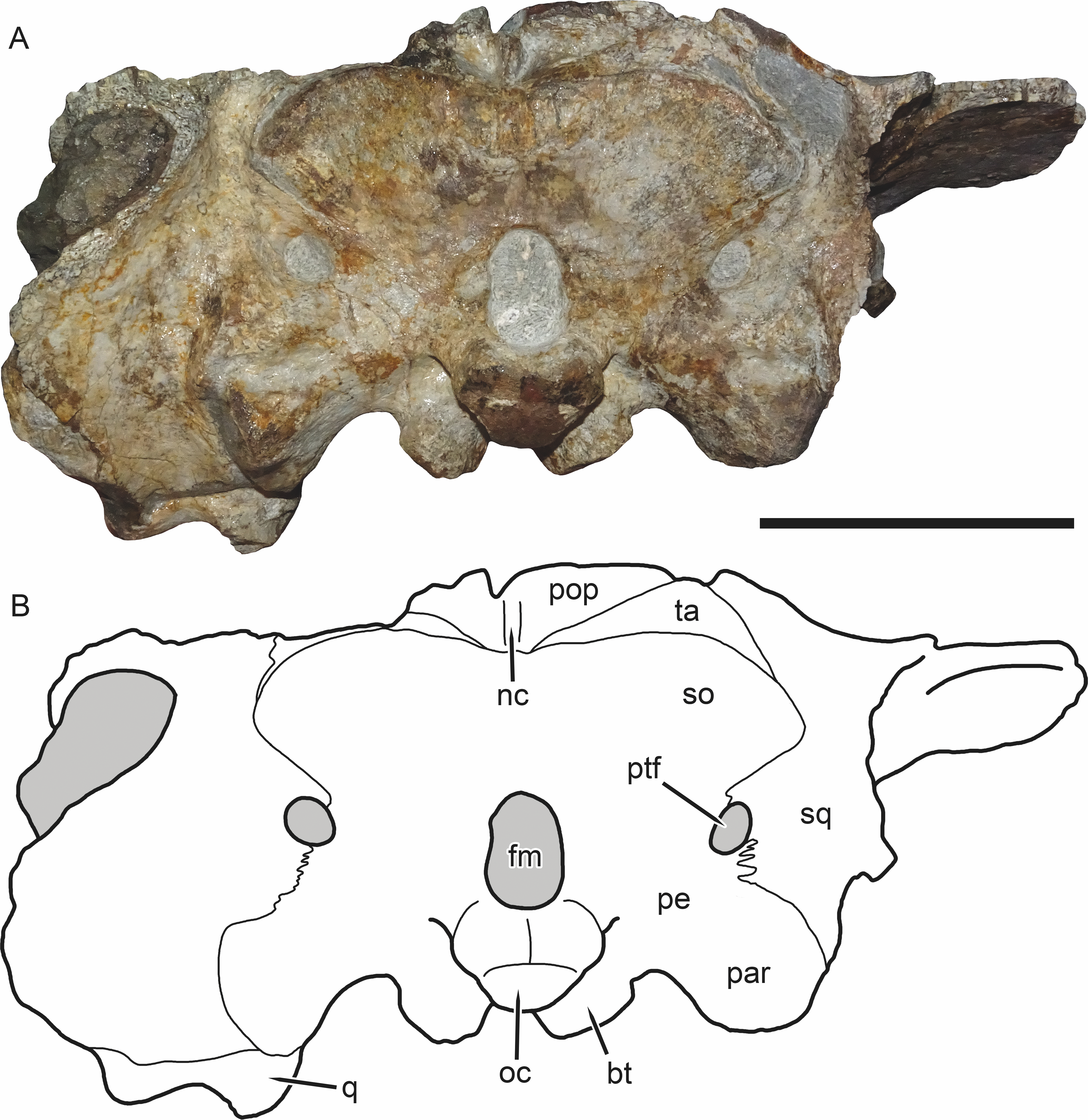{kind=link}
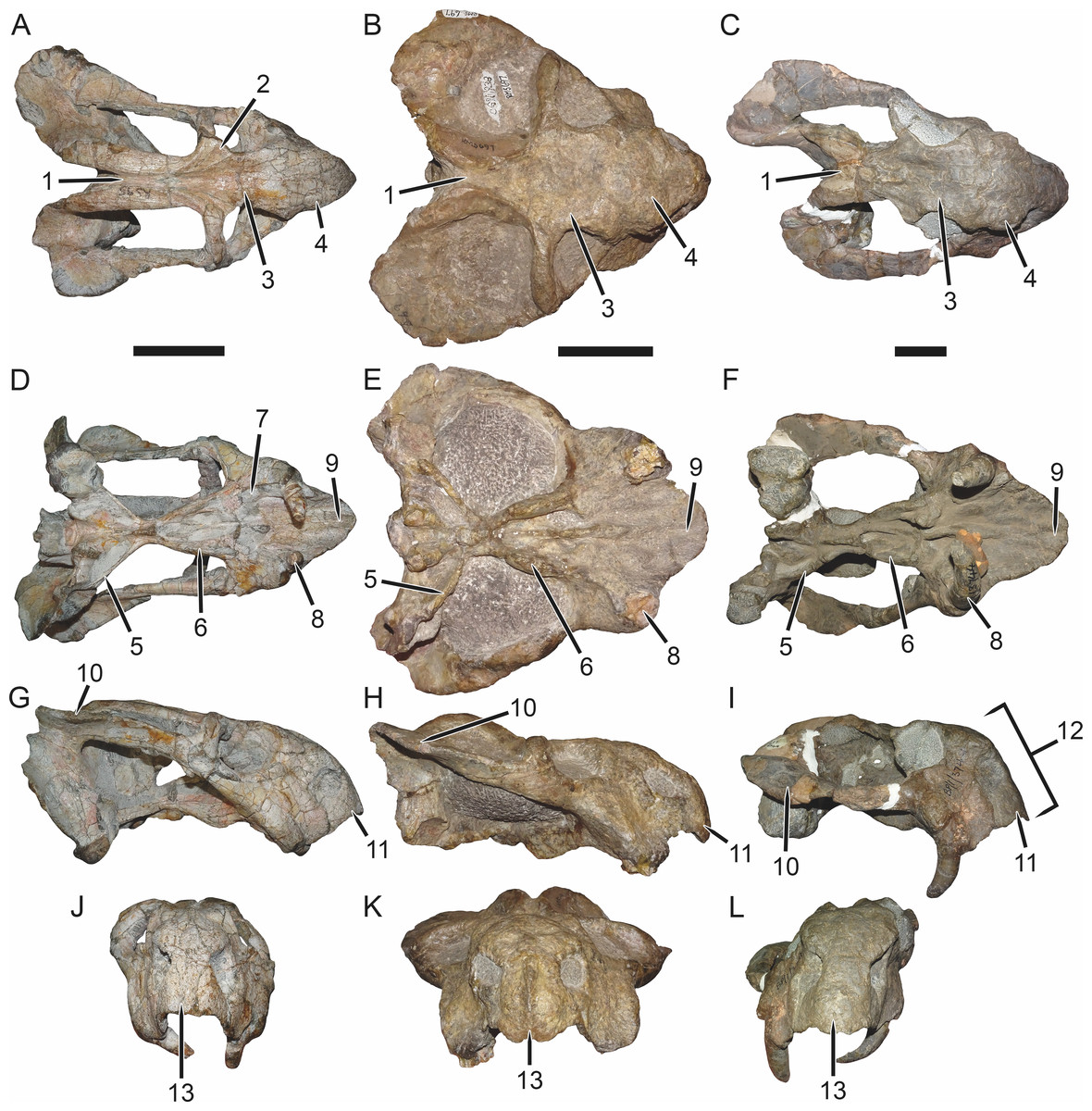
Figure 7: Comparisons of Bulbasaurus with Tropidostoma and Aulacephalodon.
Specimens: Tropidostoma dubium (SAM-PK-K11238) in (A) dorsal, (D) ventral, (G) right lateral, and (J) anterior views. Bulbasaurus phylloxyron (CGP/1/938) in (B) dorsal, (E) ventral, (H) right lateral, and (K) anterior views. Aulacephalodon bainii (BP/1/3744) in (C) dorsal, (F) ventral, (I) right lateral, and (L) anterior views. Specimen of Bulbasaurus largely undistorted; specimens of Tropidostoma and Aulacephalodon have suffered some lateral compression. Scale bars equal 5 cm; all figures of an individual specimen to scale with one another. Characters: 1, intertemporal exposure of parietal: elongate, narrow channel in Tropidostoma, ‘pinched’ but anteriorly broad in Bulbasaurus, and broad throughout but transversely expanded posteriorly in Aulacephalodon. 2, postfrontal: large and triangular in Tropidostoma, absent in Bulbasaurus and large specimens of Aulacephalodon. 3, interorbital width: narrow in Tropidostoma, relatively broad in Bulbasaurus and Aulacephalodon. 4, nasal bosses: discrete and widely-separated, transversely narrow, ovoid, smooth bosses in Tropidostoma, large, rugose bosses covering almost all of nasal surface and nearly meeting on midline in Bulbasaurus, and large, rugose, but discrete and widely-separated bosses in Aulacephalodon. 5, quadrate ramus of pterygoid: relatively thin in Tropidostoma, relatively thick in Bulbasaurus and Aulacephalodon. 6, anterior ramus of pterygoid: relatively thin in Tropidostoma, relatively thick in Bulbasaurus and Aulacephalodon. 7, postcanine teeth: present in Tropidostoma, absent in Bulbasaurus and Aulacephalodon. 8, maxillary tusk: relatively small in Tropidostoma, massive in Bulbasaurus, also massive in Aulacephalodon but only at large skull size—specimen figured here is at minimal size for large tusks in the taxon; smaller individuals have thin or just-erupting tusks. 9, depression between anterior palatal ridges: deeper than depressions lateral to ridges in Tropidostoma, of equal depth as lateral depressions in Bulbasaurus and Aulacephalodon. 10, subtemporal bar: with only a slight deflection of the dorsal edge in Tropidostoma, ‘twisted’ in Bulbasaurus, such that the medial surface of the bar becomes the dorsal surface posteriorly, and strongly ‘twisted’ in Aulacephalodon, such that the medial surface of the bar becomes the lateral surface posteriorly. 11, ‘beak’: with only weak curvature in Tropidostoma and Aulacephalodon, only extreme anterior edge with hooked tip, versus strongly ventrally-curved in Bulbasaurus, with most of premaxillary ventral margin making up hooked tip. 12, snout depth: very deep in Aulacephalodon, comparatively shallow in Tropidostoma and Bulbasaurus. 13, anterior premaxillary ridge: absent or weakly-diffuse in Tropidostoma, very discrete and well-developed in Bulbasaurus, and absent in smaller individuals of Aulacephalodon such as the one figured here (and even in the largest adults, the premaxillary ridge of Aulacephalodon is proportionally lower and more diffuse than that of Bulbasaurus).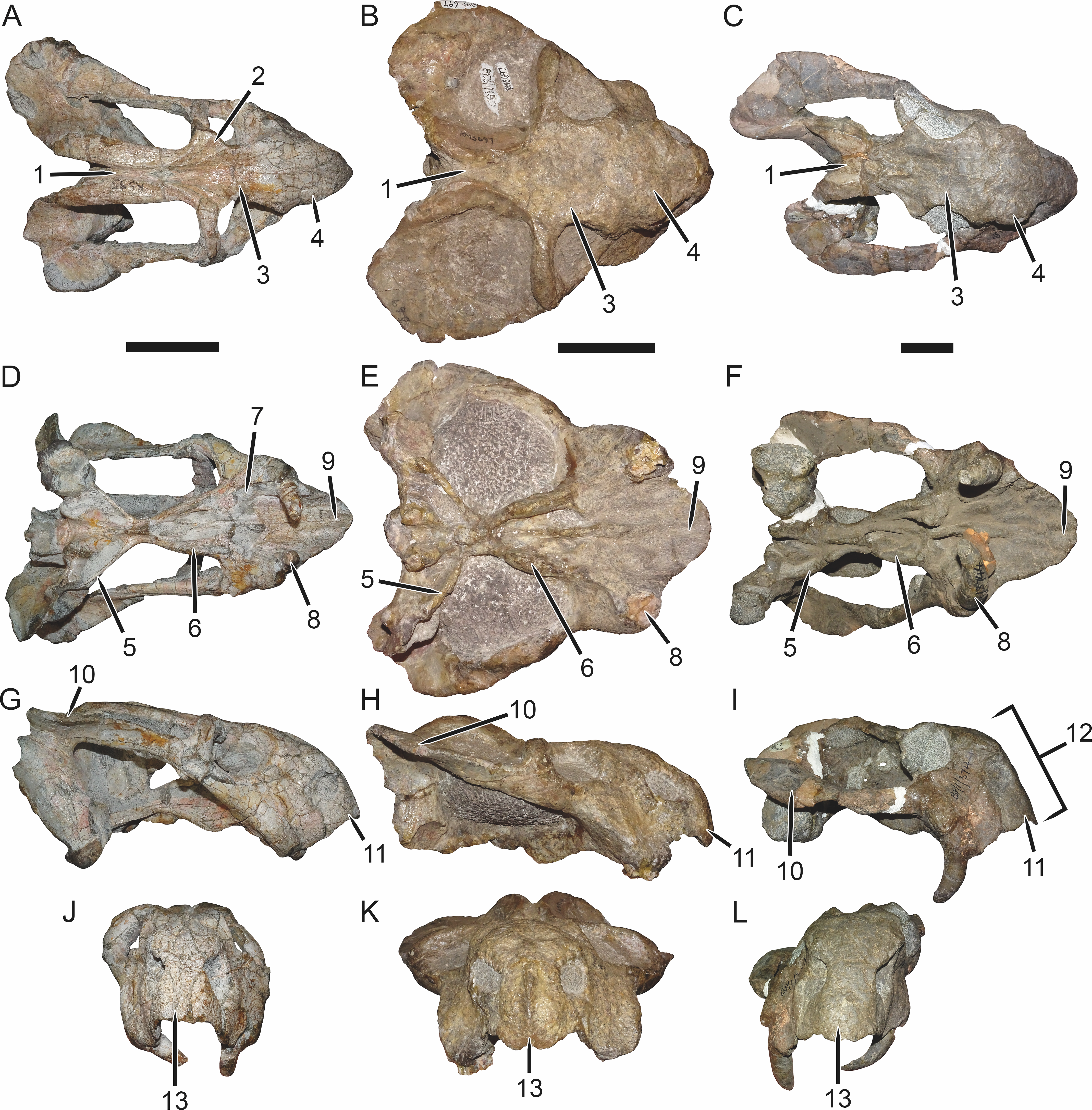{kind=link}
Referred specimens: CGP/1/938, a nearly-complete skull from Wilgerbosch Kloof, Fraserburg, Northern Cape Province, South Africa (Figs. 7B, 7E, 7H and 7K); CGP/1/949, a complete skull from Wilgerbosch Kloof, Fraserburg, Northern Cape Province, South Africa (Figs. 8–11); CGP/1/970, a complete skull, lower jaws, and partial postcranium from Blaauwkrans, Beaufort West, Western Cape Province, South Africa (Figs. 12–14); CGP/1/2263, a nearly-complete, anteroposteriorly crushed skull from an uncertain locality (Fig. 15); SAM-PK-K10106, a complete but only partially prepared, dorsoventrally crushed skull and lower jaws from Paalhuisberg, Beaufort West, Western Cape Province, South Africa; SAM-PK-K10587, a small, dorsoventrally crushed skull from Doornhoek, Beaufort West, Western Cape Province, South Africa (Fig. 16). All referred specimens are also from the Tropidostoma Assemblage Zone.
Etymology: “Leaf razor” (from the latinized Greek phyllos and xyron), referring to the slicing, inferred keratinous edge of the jaw, used in cutting plant material during feeding. A noun in apposition.
Diagnosis: Dicynodont distinguished from all taxa other than geikiids by the combination of a transverse nasofrontal ridge, extremely broad interorbital region, and twisted squamosal contribution to subtemporal bar. Distinguished from the geikiids Aulacephalodon, Pelanomodon, and Geikia by the near-confluence of the nasal bosses (covering the majority of the nasal surface), absence of prefrontal bosses, sharply curved, ‘hook’-like beak tip, tall, highly discrete premaxillary ridge, presence of massive tusks at relatively small skull size, absence of bosses or swellings on the zygomatic arch, relatively weakly twisted subtemporal bar, relatively narrow intertemporal region, and a postparietal out-of-plane with both the occipital plate and skull roof. Further distinguished from Pelanomodon and Geikia by the presence of tusks and absence of a postcaniniform crest.
Description
The following description is based primarily on the specimens SAM-PK-K11235 (the holotype), CGP/1/938, and CGP/1/949, all of which are isolated crania of similar size (Table 1). SAM-PK-K11235 is the least complete of the three and has large cracks across the intertemporal bar and snout, but was chosen as the holotype because it is the specimen that best exhibits cranial sutures. Description of the mandible and postcranium is based primarily on CGP/1/970 (one of only two Bulbasaurus specimens preserving the mandible and the only specimen preserving any postcrania). All of these specimens were previously identified as the oudenodontid Tropidostoma in collections; for this reason contrasts with Tropidostoma especially will be emphasized throughout the description, among broader comparisons with known dicynodont taxa. Comparisons with Tropidostoma dubium are based primarily on the specimens NHMUK R1662 (a topotype specimen making up part of Seeley’s (1889) original hypodigm for Tropidostoma), CGP/1/939, CGP/1/968, SAM-PK-K8603, and SAM-PK-K11238.

Figure 8: Stereopair of CGP/1/949, referred specimen of Bulbasaurus phylloxyron gen. et sp. nov., in dorsal view.
Scale bars equal 5 cm.{kind=link}
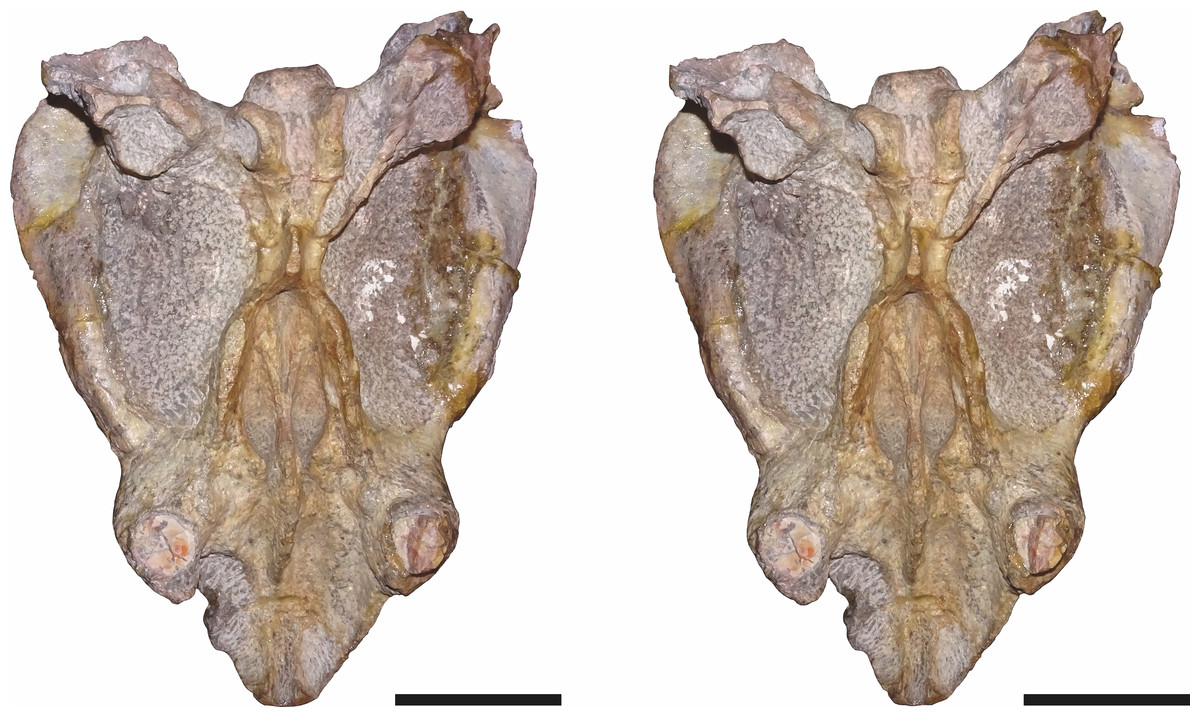
Figure 9: Stereopair of CGP/1/949, referred specimen of Bulbasaurus phylloxyron gen. et sp. nov., in ventral view.
Scale bars equal 5 cm.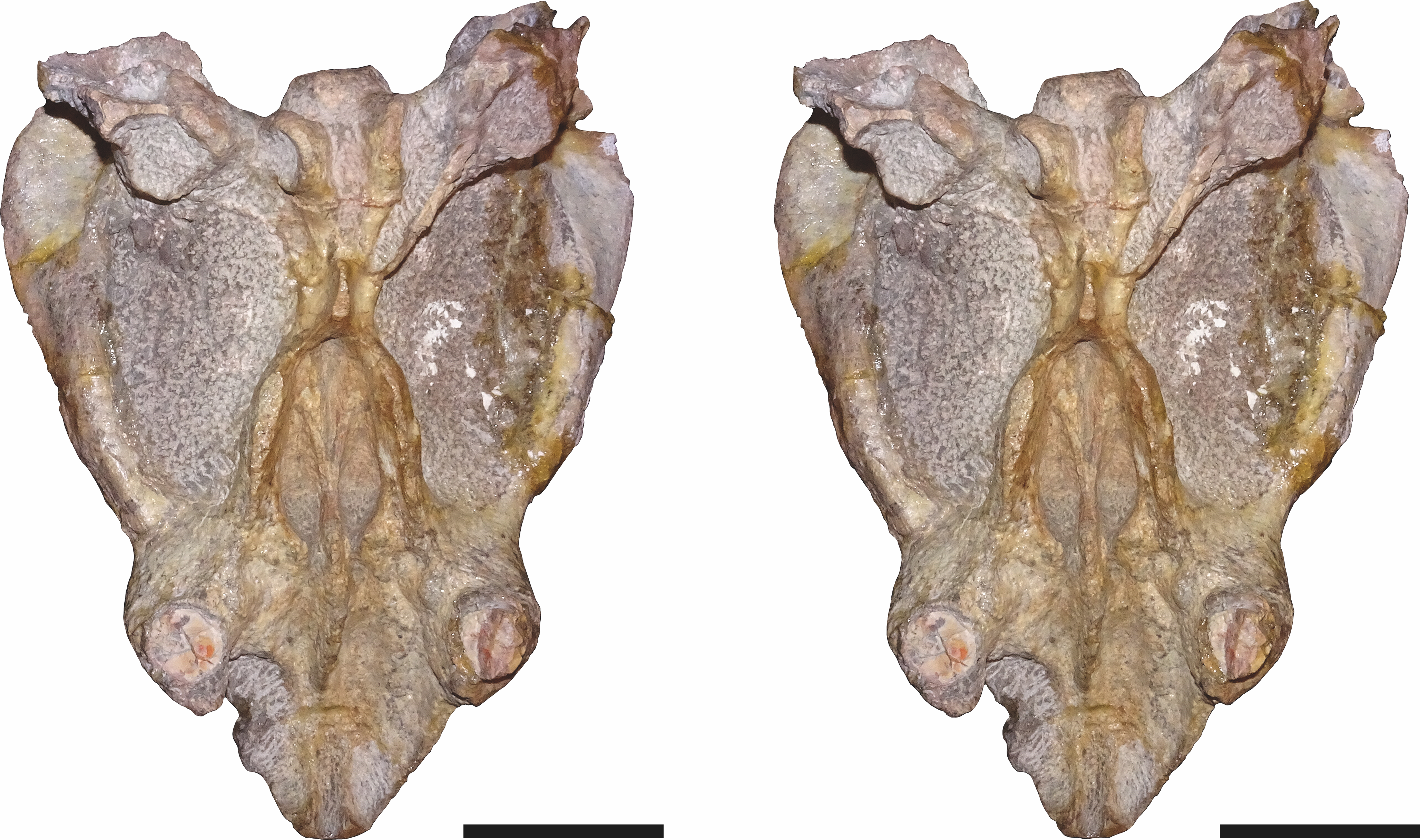{kind=link}
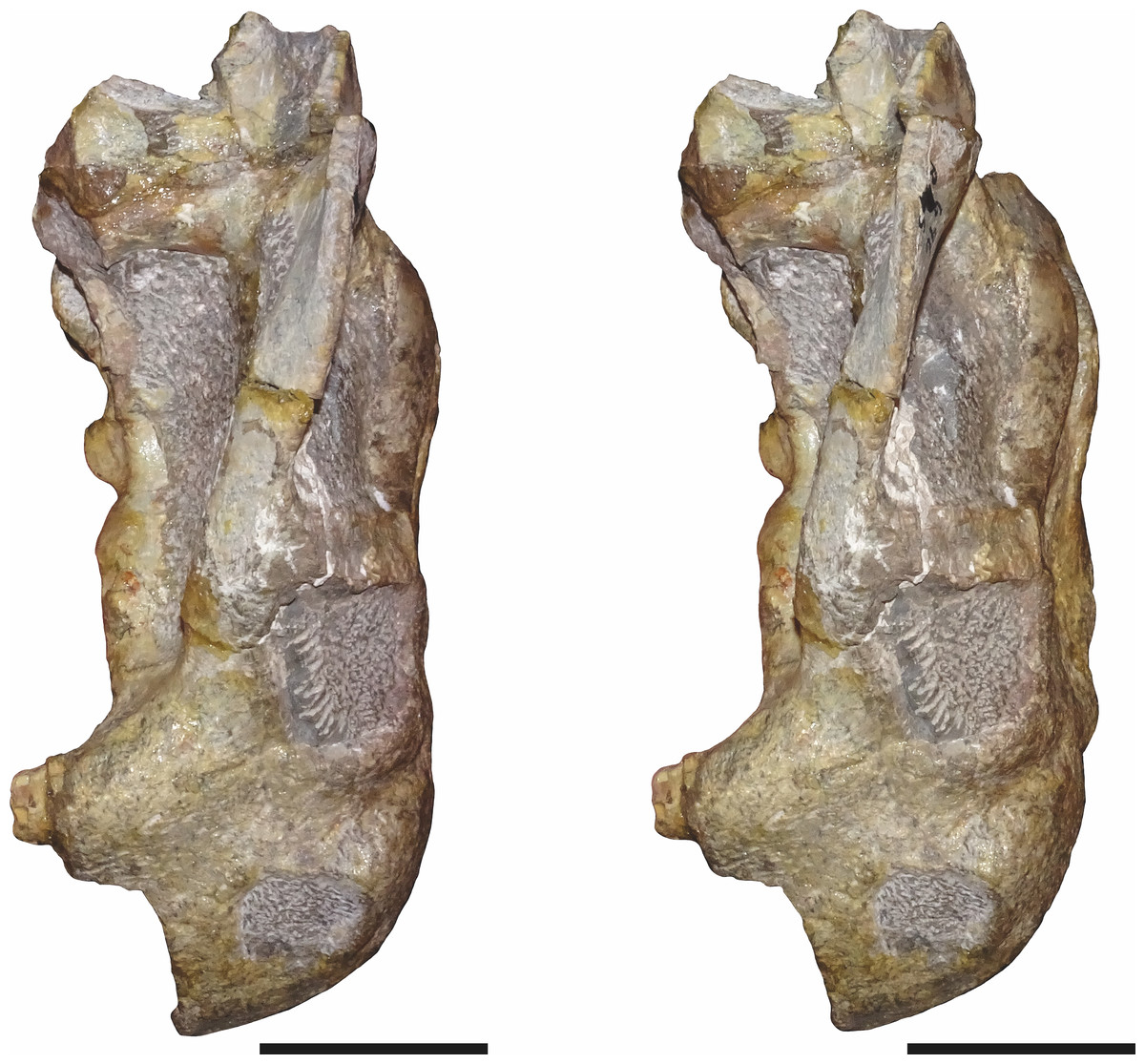
Figure 10: Stereopair of CGP/1/949, referred specimen of Bulbasaurus phylloxyron gen. et sp. nov., in right lateral view.
Scale bars equal 5 cm.{kind=link}

Figure 11: Stereopair of CGP/1/949, referred specimen of Bulbasaurus phylloxyron gen. et sp. nov., in occipital view.
Scale bars equal 5 cm.{kind=link}

Figure 12: CGP/1/970, referred specimen of Bulbasaurus phylloxyron gen. et sp. nov., in (A) dorsal and (B) left lateral views.
Postcranial elements edited out to highlight cranial morphology; see Figs. 13 and 14 for postcranium. Scale bar equals 5 cm.{kind=link}

Figure 13: Stereopair of CGP/1/970, referred specimen of Bulbasaurus phylloxyron gen. et sp. nov., in semi-dorsal view.
Scale bars equal 5 cm.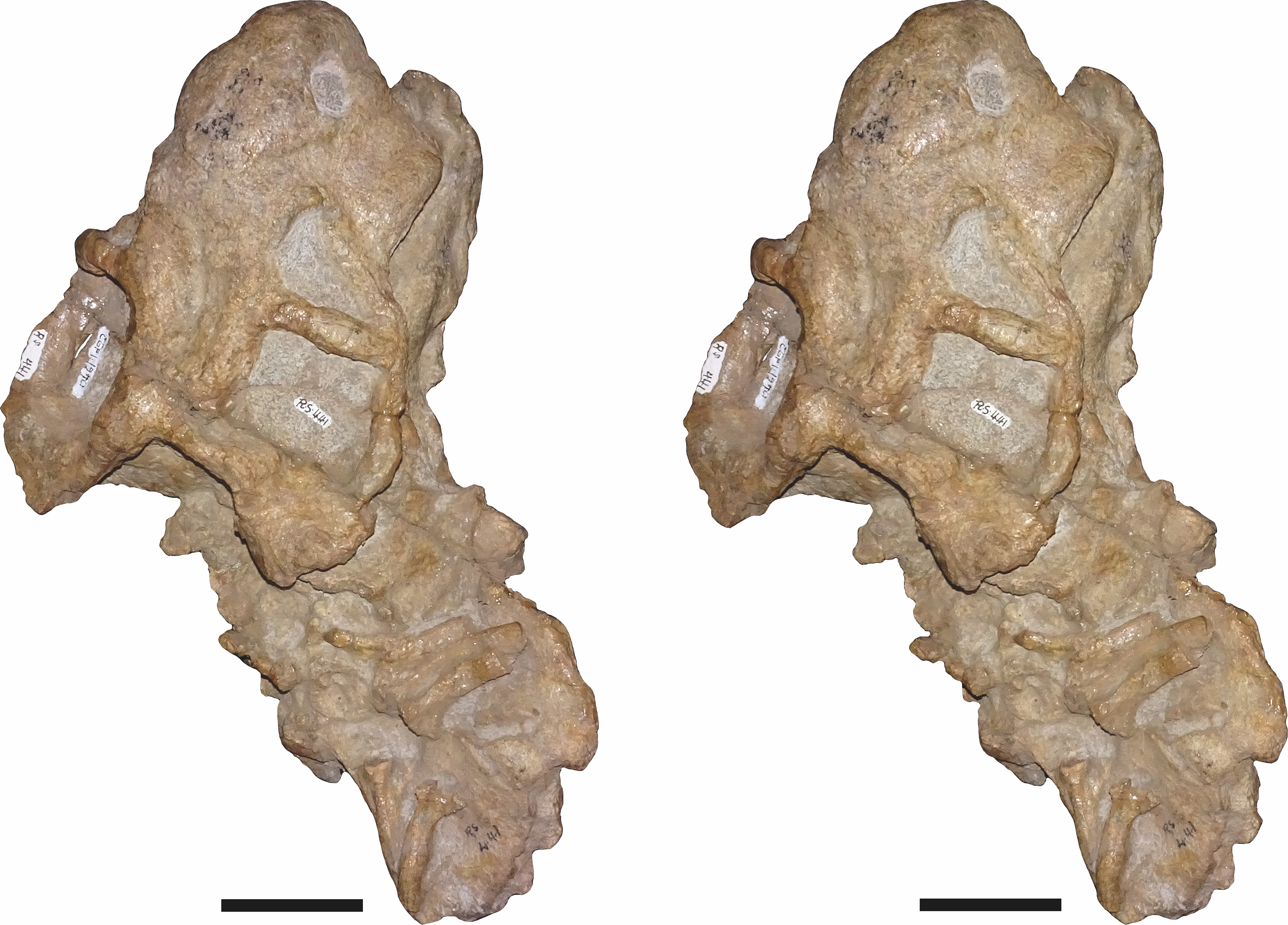{kind=link}
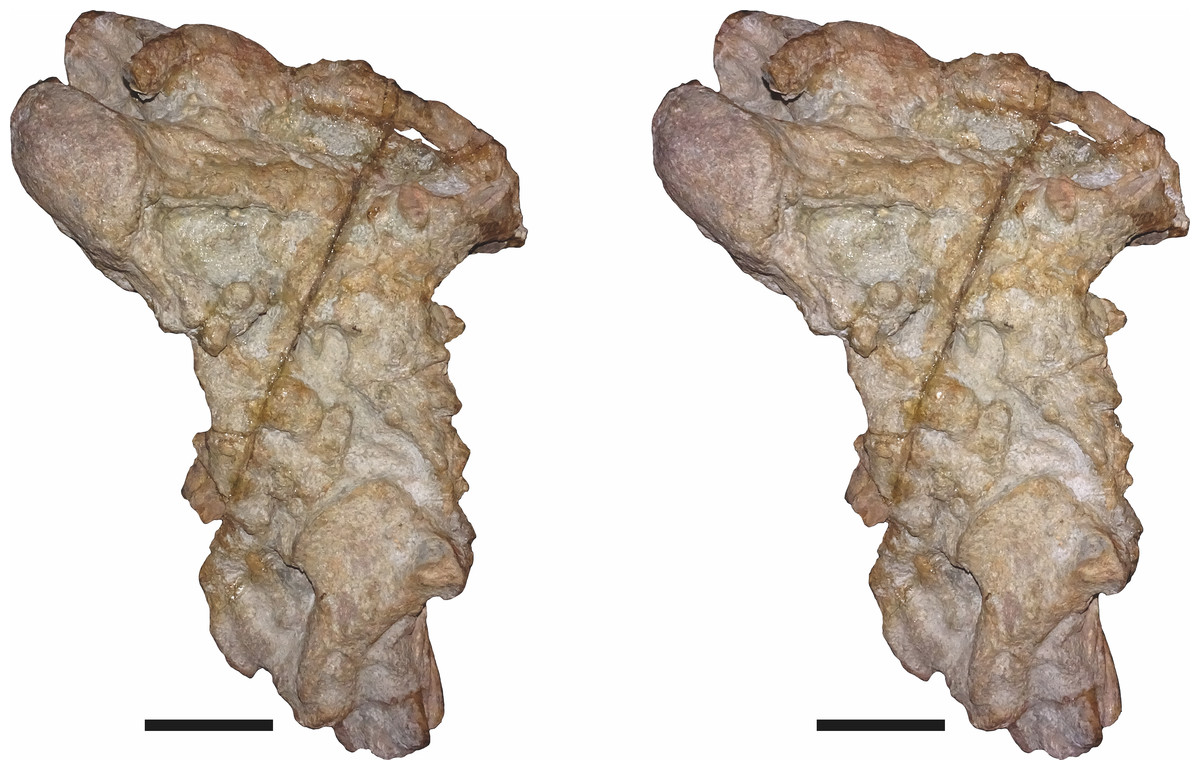
Figure 14: Stereopair of CGP/1/970, referred specimen of Bulbasaurus phylloxyron gen. et sp. nov., in ventral view.
Scale bars equal 5 cm.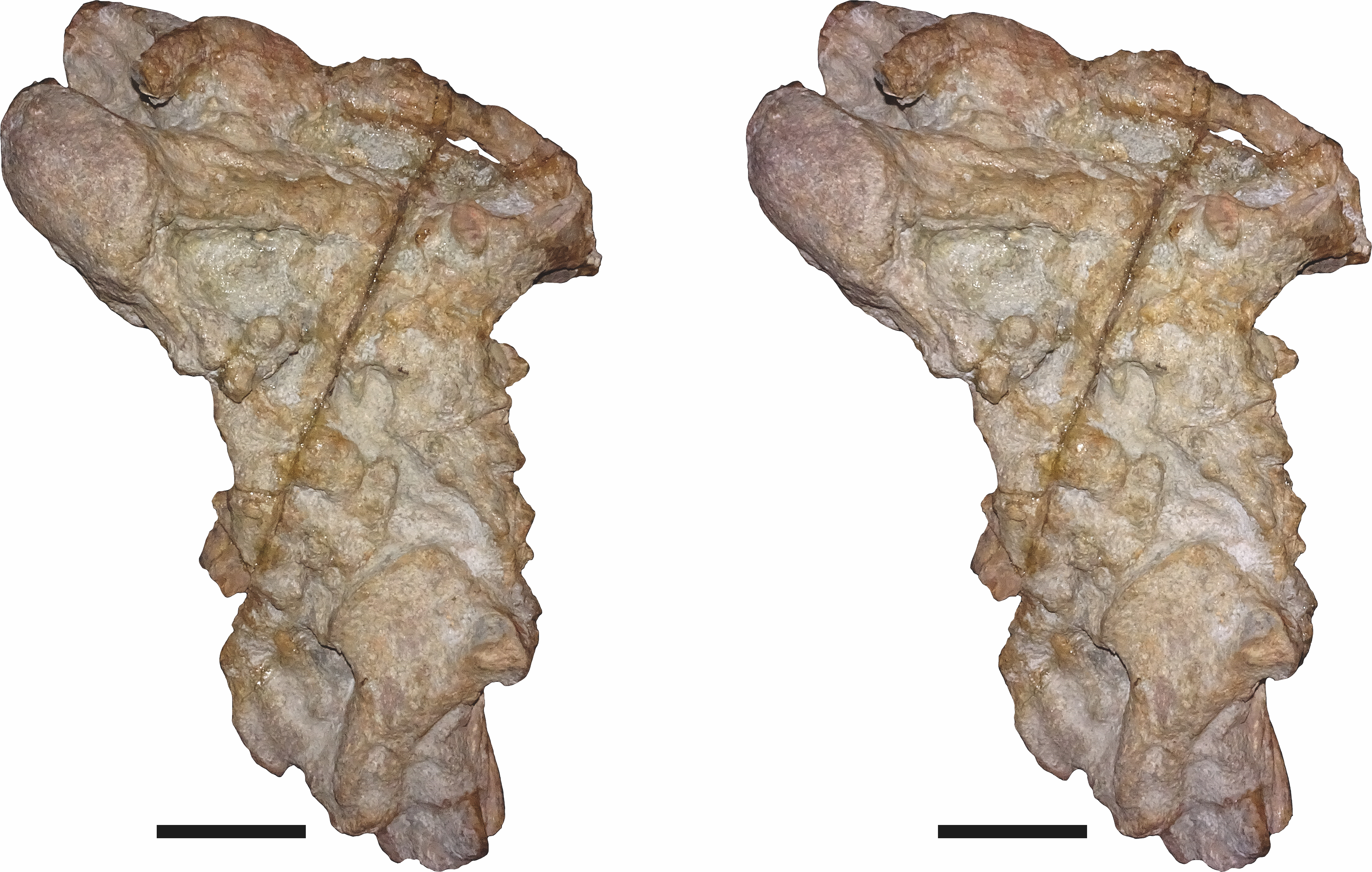{kind=link}

Figure 15: CGP/1/2263, referred specimen of Bulbasaurus phylloxyron gen. et sp. nov., in (A) dorsal, (B) right lateral, (C) occipital, (D) ventral, and (E) anterior views.
Scale bar equals 5 cm.{kind=link}
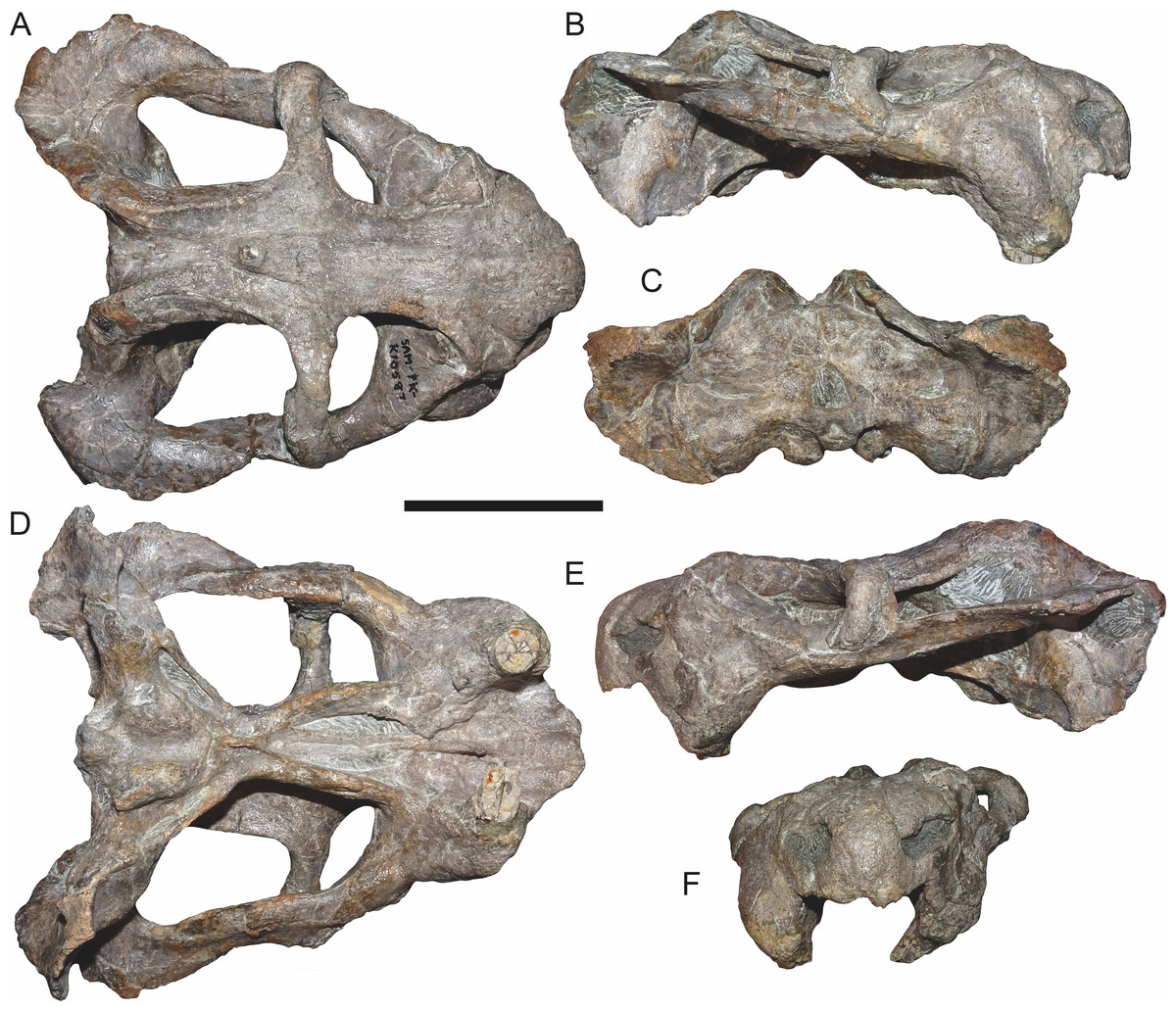
Figure 16: SAM-PK-K10587, referred specimen of Bulbasaurus phylloxyron gen. et sp. nov., in (A) dorsal, (B) right lateral, (C) occipital, (D) ventral, (E) left lateral, and (F) anterior views.
Scale bar equals 5 cm.{kind=link}
The premaxillae of Bulbasaurus phylloxyron are fused to form a single median element, as in most dicynodonts (King, 1988; Kammerer & Angielczyk, 2009). On the dorsal skull surface, the premaxilla forms the anterior tip of the ‘beak’; palatally it forms a broad plate making up most of the secondary palate (Figs. 2–5). The tip of the ‘beak’ is sharply deflected in Bulbasaurus (Figs. 3, 7H); although not to the extreme degree of the dicynodontoid Dinanomodon (Kammerer, Angielczyk & Fröbisch, 2011), it is substantially more ‘hook’-like than in Tropidostoma and other cryptodonts (Figs. 7G and 7I). The anterior face of the premaxilla is flattened and bears a highly discrete median ridge with weak depressions on either side (Figs. 2A and 7K). In Tropidostoma, although there is usually a median ridge, it is relatively weak and diffuse (Fig. 7J). A well-developed premaxillary ridge is also present in large specimens of Aulacephalodon, although it is generally broader and more diffuse at edge than the tall, sharp ridge of Bulbasaurus. Posterodorsal to the median ridge, the premaxilla becomes a sharply tapering structure (the ascending process) narrowly extending between the nasal bosses. The posterior extent of the ascending process varies between specimens: it is remarkably long in the holotype, nearly reaching the level of the nasofrontal suture (Fig. 4B), but is relatively short in CGP/1/2263 (Fig. 15A) (although it also nearly reaches the nasofrontal suture in that specimen because of an elongate anterior process of the frontal, which is absent in the holotype). Laterally, the premaxilla contacts the maxilla below the septomaxillary footplate of the external naris (Figs. 2A and 3B). The premaxilla is roughly pentagonal in outline in palatal view (Fig. 5B). Well-developed, paired anterior palatal ridges and a single posterior median ridge are present. The paired anterior palatal ridges are relatively close together, as in Aulacephalodon and Pelanomodon (Kammerer, Angielczyk & Fröbisch, 2015a). Elongate depressions are present lateral to these ridges, and an additional, median depression is present between them. In Bulbasaurus, these three depressions are all of equal depth, as in other geikiids (Fig. 7F) but unlike Tropidostoma in which the median depression is substantially deeper than the lateral ones (Fig. 7D). The median ridge is both taller and transversely broader than the anterior ridges. It originates as a low, narrow structure at the posterior edge of the paired anterior ridges, but is not confluent with them (Figs. 5 and 9). It increases in height and breadth posteriorly, with maximal size achieved immediately anterior to the palatine pads. This ridge continues into the interpterygoid region, where it becomes formed by the vomer. Immediately lateral to the median palatal ridge the premaxillary surface is only weakly sloping; there is not a distinct, elongate depression extending along the side of the ridge. However, there are paired premaxillary depressions further laterally, in the region around the anterior terminus of the median ridge (Fig. 5B). Lateral to these depressions are low, rugose, somewhat arcuate ridges near the bases of the caniniform processes. Posteriorly, each of these ridges terminates at a confluence with the anteromedial margin of the palatine pad. Immediately lateral to this confluence is a small but discrete circular fossa at the anterior midpoint of the palatine pad, representing the lateral palatal foramen.
The septomaxilla is a small element largely confined within the external naris (Fig. 2A), but also making up the ventral rim of the naris in the form of a footplate. It is similar in morphology to that of other cryptodonts.
The maxilla is a large bone making up the ventrolateral surface of the snout (Fig. 3B). It bears a well-developed caniniform process housing a massive tusk. The tusks of Bulbasaurus are enormous proportional to skull size; in SAM-PK-K11235 the tusk diameter is 1.9 cm on a skull of 14.0 cm dorsal length (see also Table 2). Tusks in Tropidostoma are consistently proportionally smaller than those of Bulbasaurus, and absolutely smaller in all but the largest known Tropidostoma specimens (Table 3). The mean relative tusk diameter (RTD; measured as the ratio of tusk diameter to dorsal skull length) of Bulbasaurus is 0.13 (Table 2); by comparison, the mean RTD of Tropidostoma is only 0.06 (Table 3). Tusk proportions similar to those of Bulbasaurus are otherwise known only in Aulacephalodon among cryptodonts. However, Bulbasaurus is remarkable in having such large tusks at relatively small skull size. Aulacephalodon skulls of comparable size to specimens of Bulbasaurus are usually considered juveniles (Tollman, Grine & Hahn, 1980), and have the small, often still-erupting tusks typical of young dicynodonts. Aulacephalodon specimens only exhibit adult tusk proportions (i.e., those with RTD >0.10) in skulls greater than 20 cm in length (Table 4), and even these adults tend to have proportionally smaller (mean RTD 0.11) tusks than Bulbasaurus (although some individuals attain equivalent proportions). Because of the large size of the tusk root in Bulbasaurus, the caniniform process prominently bulges outwards along its lateral surface. The lateral surface of the maxilla is covered in tiny foramina and scattered larger pits. The rugose surface of the entire snout is suggestive of a keratinous covering, as has been argued to be broadly present in dicynodonts (King, 1988; Kammerer, Angielczyk & Fröbisch, 2015b). Dorsally, the maxilla extends between the lacrimal and external naris, before contacting the nasal and anteroventral tip of the prefrontal at its dorsal terminus (Fig. 3B). Posteriorly, it forms an elongate process contributing to the zygomatic arch, underlying first the jugal and then the squamosal before terminating beneath the postorbital bar. The posteroventral face of the caniniform process has a noticeably smoother bone texture than the lateral surface of the maxilla, broken only by two large foramina. No postcaniniform crest is present, unlike all cryptodonts other than Aulacephalodon (although in juvenile Aulacephalodon of comparable size to Bulbasaurus, e.g., BSPG 1934-VIII-516, a postcaniniform crest is present). Posteriorly, there is a straight suture with the jugal and, more medially, an interdigitated suture with the ectopterygoid and palatine (Fig. 5B). A labial fossa is present in this region (Fig. 15D), but the maxilla does not contribute to its border, which is formed by the jugal, ectopterygoid, and palatine.
The nasal forms the roof of the snout and bears a large boss. Paired nasal bosses are present in all known cryptodonts (Kammerer & Angielczyk, 2009) and come in various shapes and sizes within the clade. The nasal bosses of Tropidostoma are ovoid and proportionally transversely narrow (Fig. 7A), similar to those of Oudenodon and to a lesser extent Rhachiocephalus. Additionally, they are clearly separate structures, with unornamented nasal surfaces and a broad, triangular dorsal process of the premaxilla intervening between them. In Bulbasaurus, by contrast, the nasal bosses are massive and nearly confluent, with only a narrow strip of premaxilla separating them for most of their length (Figs. 2, 4, 7B and 8). Furthermore the nasal bosses of Bulbasaurus are notably rugose, whereas those of Tropidostoma, although bearing some pits, have a smoother bone surface. The nasal bosses of Aulacephalodon and Pelanomodon are also massive and frequently rugose, but unlike Bulbasaurus are also separated by broad spans of unornamented nasal (Fig. 7C). A distinct dorsoventral depression extending across the nasal, prefrontal, and maxilla separates the nasal boss from the orbital rim (Fig. 10), as in almost all other cryptodonts. Only in Odontocyclops is the nasal boss completely confluent with a prefrontal boss (Angielczyk, 2002). Posterior to the terminus of the ascending process of the premaxilla, the nasals have a short midline suture (Figs. 4B and 15A). This suture is situated in a narrow depression, in the same position where some other dicynodonts have fontanelles during development or neomorphic snout bones (Jasinoski et al., 2014; Kammerer, Angielczyk & Fröbisch, 2015a). Posteriorly, the nasal has a nearly straight but highly interdigitated suture with the frontal. In the holotype this interdigitation is dense but the individual processes are relatively short (Fig. 4), whereas in CGP/1/2263 there is a combination of dense interdigitation and a few large processes (notably an anterior process of the frontals that nearly extends between the nasal bosses; Fig. 15A). This suture is weakly but distinctly raised relative to the surrounding bone. Although not as well-developed as the tall nasofrontal ridge of Aulacephalodon and Pelanomodon, this raised suture is quite unlike the condition in Tropidostoma, in which there is no difference in skull height across the nasofrontal border.
The lacrimal is a narrow, curved bone that on the snout surface only forms the middle part of the anterior orbital margin (Fig. 3B). The lacrimal contribution to the orbital margin bears the best-developed portion of the orbital ridge, which continues onto the anterior portions of the jugal and prefrontal, but is absent around the rest of the orbital rim. Along its anteroventral margin, the lacrimal is excluded from contacting the naris by the dorsal process of the maxilla. Along its anterodorsal margin it has a variable extent between individuals, sometimes contacting the nasal (Fig. 15B) and sometimes being excluded from the nasal by a thin strip of prefrontal (Fig. 3B). Within the orbit, the lacrimal is perforated by a single, large lacrimal foramen.
The prefrontal is also a small bone that is largely limited to the anterior orbital margin (Figs. 3B–4B). Although its anterodorsal surface is somewhat thickened and convex, no distinct prefrontal boss is present (Figs. 4 and 8), unlike the majority of cryptodonts. Indeed, the lateral surface of the prefrontal in front of the orbit is less protruding than that of the lacrimal, as mentioned above.
| Specimens | Basal skull length (cm) | Dorsal skull length (cm) | Snout length (cm) | Interorbital least width (cm) | Anterior intertemporal width (cm) | Posterior intertemporal width (cm) | Temporal fenestra length (cm) |
|---|---|---|---|---|---|---|---|
| CGP/1/938 | 15.1 | 13.4 | 4.2 | 4.8 | 3.9 | 1.6 | 7.9 |
| CGP/1/949 | 16.4 | 13.1 | 4.5 | 4.2 | 4.4 | 3.9 | 7.0 |
| CGP/1/970 | N/A | 16.0 | 6.6 | 5.2 | 5.3 | 0.6 | 9.0 |
| CGP/1/2263 | 11.1 | 10.9 | 3.5 | 2.4 | 3.5 | 4.0 | 6.7 |
| SAM-PK-K10106 | N/A | 13.4 | 5.0 | 4.8 | 4.2 | 1.1 | 8.7 |
| SAM-PK-K10587 | 11.1 | 10.4 | 2.7 | 2.2 | 3.6 | 3.5 | 6.1 |
| SAM-PK-K11235 | 16.9 | 14.0 | 5.2 | 4.4 | 4.0 | 0.6 | 9.6 |
| Mean (all specimens) | 14.1 | 13.0 | 4.5 | 4.0 | 4.1 | 2.2 | 7.9 |
| Mean (excluding CGP/1/2263 and SAM-PK-K10587) | 16.1 | 14.0 | 5.1 | 4.7 | 4.4 | 1.6 | 8.4 |
| Standard deviation (all specimens) | 2.8 | 1.9 | 1.3 | 1.2 | 0.6 | 1.6 | 1.3 |
| Specimens | Interpterygoid vacuity length (cm) | Median pterygoid plate width (cm) | Angle of quadrate ramus (left) | Angle of quadrate ramus (right) | Angle between squamosal rami | Dentary symphysis height (cm) | Dentary ramus height (cm) |
|---|---|---|---|---|---|---|---|
| CGP/1/938 | 0.8 | 1.7 | 53.0 | 63.0 | 150.0 | N/A | N/A |
| CGP/1/949 | 0.9 | 1.7 | N/A | 40.0 | 149.0 | N/A | N/A |
| CGP/1/970 | N/A | N/A | N/A | N/A | N/A | 9.8 | 6.3 |
| CGP/1/2263 | N/A | N/A | 69.0 | 66.0 | 144.0 | N/A | N/A |
| SAM-PK-K10106 | N/A | N/A | N/A | N/A | N/A | N/A | N/A |
| SAM-PK-K10587 | 0.6 | 1.4 | 44.0 | 52.0 | 150.0 | N/A | N/A |
| SAM-PK-K11235 | 0.8 | 1.8 | 58.0 | 56.0 | 146.0 | N/A | N/A |
| Mean (all specimens) | 0.8 | 1.7 | 56.0 | 55.4 | 147.8 | 9.8 | 6.3 |
| Mean (excluding CGP/1/2263 and SAM-PK-K10587) | 0.8 | 1.7 | 55.5 | 53.0 | 148.3 | 9.8 | 6.3 |
| Standard deviation (all specimens) | 0.1 | 0.2 | 10.4 | 10.2 | 2.7 | N/A | N/A |
| Specimens | Dorsal skull length (DSL) (cm) | Left tusk diameter (cm) | Right tusk diameter (cm) | Tusk diameter mean (TDM) (cm) | Relative tusk diameter (TDM/DSL) |
|---|---|---|---|---|---|
| CGP/1/938 | 13.4 | 1.9 | 2.2 | 2.1 | 0.15 |
| CGP/1/949 | 13.1 | 1.4 | 1.7 | 1.6 | 0.12 |
| CGP/1/970 | 16.0 | 2.2 | N/A | 2.2 | 0.14 |
| CGP/1/2263 | 10.9 | 1.2 | 1.3 | 1.3 | 0.12 |
| SAM-PK-K10106 | 13.4 | 1.8 | 1.7 | 1.8 | 0.13 |
| SAM-PK-K10587 | 10.4 | 1.2 | 1.3 | 1.3 | 0.12 |
| SAM-PK-K11235 | 14.0 | 2.0 | 1.8 | 1.9 | 0.13 |
| Mean | 13.0 | 1.7 | 1.7 | 1.7 | 0.13 |
| Standard Deviation | 1.9 | 0.4 | 0.3 | 0.4 | 0.01 |
| Specimens | Dorsal skull length (DSL) (cm) | Left tusk diameter (cm) | Right tusk diameter (cm) | Tusk diameter mean (TDM) (cm) | Relative tusk diameter (TDM/DSL) |
|---|---|---|---|---|---|
| CGP/1/930 | 13.4 | 1.1 | 1.1 | 1.1 | 0.08 |
| CGP/1/2173 | 12.1 | 0.5 | 0.6 | 0.5 | 0.04 |
| CGS CM86-573 | 17.0 | 0.9 | 0.9 | 0.9 | 0.05 |
| CGS F7 | 19.1 | 1.1 | 0.7 | 0.9 | 0.05 |
| CGS F11 | 19.9 | 0.5 | 1.4 | 1.0 | 0.05 |
| CGS R98 | 13.1 | 0.9 | 0.9 | 0.9 | 0.07 |
| CGS RMS155 | 14.9 | 0.9 | 1.3 | 1.1 | 0.07 |
| CGS RMS213 | 20.9 | 1.6 | 1.8 | 1.7 | 0.08 |
| CGS RMS244 | 14.0 | 0.5 | N/A | 0.5 | 0.03 |
| CGS RMS631 | 18.8 | 0.9 | 0.8 | 0.8 | 0.04 |
| CGS RS327 | 22.4 | 1.9 | 1.5 | 1.7 | 0.08 |
| CGS RS538 | 13.9 | 0.6 | 0.8 | 0.7 | 0.05 |
| NHMUK R1662 | 25.3 | 2.1 | 1.5 | 1.8 | 0.07 |
| NHMUK R4048 | 19.1 | 1.7 | 1.7 | 1.7 | 0.09 |
| SAM-PK-K6742 | 23.4 | N/A | 1.4 | 1.4 | 0.06 |
| SAM-PK-K6808 | 18.0 | 0.5 | N/A | 0.5 | 0.03 |
| SAM-PK-K6940 | 14.1 | 0.5 | 0.3 | 0.4 | 0.03 |
| SAM-PK-K8603 | 17.7 | 1.1 | 1.0 | 1.1 | 0.06 |
| SAM-PK-K8639 | 16.2 | 0.9 | 0.4 | 0.7 | 0.04 |
| SAM-PK-K10681 | 16.8 | 1.7 | 1.5 | 1.6 | 0.09 |
| SAM-PK-K11015 | 13.0 | 1.1 | 1.2 | 1.1 | 0.09 |
| SAM-PK-K11183 | 22.3 | 1.5 | 1.3 | 1.4 | 0.06 |
| SAM-PK-K11238 | 13.6 | 1.0 | 1.0 | 1.0 | 0.07 |
| SAM-PK-K11255 | 21.8 | 2.3 | 1.0 | 1.6 | 0.08 |
| Mean | 17.5 | 1.1 | 1.1 | 1.1 | 0.06 |
| Standard deviation | 3.8 | 0.5 | 0.4 | 0.4 | 0.02 |
| Specimens | Dorsal skull length (DSL) (cm) | Left tusk diameter (cm) | Right tusk diameter (cm) | Left/right tusk diameter mean (TDM) (cm) | Relative tusk diameter (TDM/DSL) |
|---|---|---|---|---|---|
| BP/1/300 | 18.2 | 0.6 | 0.8 | 0.7 | 0.04 |
| BP/1/304 | 40.0 | 4.4 | 3.9 | 4.2 | 0.10 |
| BP/1/2460 | 34.2 | 3.8 | 3.0 | 3.4 | 0.10 |
| BP/1/2983 | 36.9 | 4.8 | 3.9 | 4.3 | 0.12 |
| BP/1/4087 | 25.3 | 3.5 | 3.1 | 3.3 | 0.13 |
| BSPG 1934-VIII-516 | 14.5 | 0.9 | 0.9 | 0.9 | 0.06 |
| CGP/1/268 | 22.6 | 2.5 | 2.4 | 2.5 | 0.11 |
| CGP/1/657 | 26.4 | 3.0 | 3.0 | 3.0 | 0.11 |
| CGP/1/740 | 28.7 | 3.4 | 2.8 | 3.1 | 0.11 |
| CGS S6 | 24.5 | 2.8 | 3.3 | 3.1 | 0.12 |
| SAM-PK-3423 | 22.7 | 2.7 | 2.8 | 2.7 | 0.12 |
| SAM-PK-5862 | 15.2 | 0.9 | 0.8 | 0.9 | 0.06 |
| SAM-PK-10021 | 33.9 | 3.9 | 3.8 | 3.8 | 0.11 |
| SAM-PK-10048 | 18.6 | 1.3 | 1.3 | 1.3 | 0.07 |
| SAM-PK-10673 | 25.1 | 2.7 | 2.4 | 2.6 | 0.10 |
| USNM 24621 | 37.0 | 3.2 | 4.3 | 3.7 | 0.10 |
| Mean (all specimens) | 26.5 | 2.8 | 2.7 | 2.7 | 0.10 |
| Mean (specimens with >20 cm skull length) | 29.8 | 3.4 | 3.2 | 3.3 | 0.11 |
| Standard deviation | 8.0 | 1.3 | 1.1 | 1.2 | 0.03 |
The jugal is an elongate bone that is primarily part of the zygoma (Fig. 4B). It has a short facial exposure above the posterior portion of the caniniform process and makes up the ventral wall of the orbit (Fig. 3B). As previously described, it also borders the ventral surface of the maxilla, behind the caniniform process (Fig. 5B), where it partially surrounds the labial fossa. From here, the jugal curves posterolaterally, forming the ventromedial edge of the zygoma until reaching the end of the temporal fenestra. It also has a short contribution to the dorsal surface of the subtemporal bar, immediately behind the postorbital bar, and also forms part of the posteromedial face of the postorbital bar. It is generally similar in morphology to that of other cryptodonts (although Pelanomodon is aberrant in having a jugal boss as part of its contribution to the postorbital bar; Kammerer, Angielczyk & Fröbisch, 2015a).
The zygomatic ramus of the squamosal tapers anteriorly, separating the maxilla and jugal below the postorbital bar before terminating below the orbital midlength (Fig. 3B). The squamosal is prominently twisted in the subtemporal bar, such that what was its lateral surface anteriorly becomes ventral surface posteriorly. Some deflection of the edge of the squamosal in the posterior part of the subtemporal bar is typical in cryptodonts (including Tropidostoma; Fig. 7G), but in the large geikiids this is taken to an extreme, with complete reversal of the lateral and medial faces of the zygoma along its length (Fig. 7I). Bulbasaurus exhibits an intermediate condition, with some twisting of the zygoma (Fig. 7H) but not to the degree seen in Aulacephalodon and Pelanomodon. The squamosal flares broadly posterolaterally around the temporal fenestra before curving inwards to form a short intertemporal ramus immediately behind the postorbital (Fig. 4B). Posteriorly, the squamosal is a major contributor to the occipital plate, bordering the tabular and fused supraoccipital-periotic element laterally (Fig. 6B). Medial extensions of the squamosal surround the lateral half of the post-temporal fenestra, as in several other cryptodont taxa (Aulacephalodon, Pelanomodon, Oudenodon (Kammerer, Angielczyk & Fröbisch, 2015a), and Tropidostoma (C Kammerer, pers. obs., 2016)) A mediolaterally deep, dorsoventrally narrow notch is present between the zygomatic and quadrate rami of the squamosal (Fig. 11).
The frontal is largely restricted to the interorbital region in Bulbasaurus (Fig. 4B). It has a rugose edge where it forms part of the dorsal margin of the orbit, and several deep pits on its dorsal surface. The frontal curves downwards posteromedially, forming a deep median depression with the preparietal between the postorbital bars, anterior to the pineal foramen (Figs. 4 and 8). A depression in this part of the skull is sometimes weakly developed in Tropidostoma and Oudenodon, but is absent in other geikiids. This is probably related to the general broadening and flattening of the intertemporal region in these taxa relative to most other bidentalians. Posterolaterally, the border between the frontal and postorbital forms a sharp ridge. A distinct postfrontal appears to be absent in Bulbasaurus(Fig. 4B). The postfrontal is a large, triangular element at the posterodorsal margin of the orbit in Tropidostoma (Fig. 7A) and is usually narrower but still distinct in Oudenodon. The postfrontal is not discernible in adult geikiids such as Aulacephalodon, although it appears to be present in some juvenile specimens (e.g., BSPG 1934-VIII-516), suggesting that it fuses with the postorbital during growth. The absence of a distinct postfrontal in Bulbasaurus is further evidence of ‘adult’ geikiid characters being present at small skull size in this taxon.
The postorbital makes up the majority of the postorbital bar and the medial edge of the temporal fenestra (Figs. 3B, 4B and 8). The postorbital bar is typically thick in cryptodonts, but is particularly robust in geikiids, Bulbasaurus included. Unlike other geikiids, however, the postorbital bar of Bulbasaurus does not bear any bosses or dorsoventral ridges. The posterior ramus of the postorbital has a very steep lateral face, with a sharp break in slope between the postorbital laterally and the skull roof (frontal and parietal) medially. The lateral face of the postorbital is distinctly concave and would have served as the attachment site for jaw musculature. The posterior rami of the postorbitals converge posteriorly, creating a ‘pinched’ intertemporal bar. The degree of this convergence varies between specimens; in SAM-PK-K11235, the postorbitals almost completely overlap the parietals posteriorly, with only a narrow strip of parietal exposed between them (Fig. 4), whereas in CGP/1/938 and CGP/1/949, a trough-like median exposure of the skull roof is retained in the posterior intertemporal bar (Figs. 7B and 8). This style of variability is common in Tropidostoma and Oudenodon; in adult specimens of Aulacephalodon and Pelanomodon the parietal always remains broadly exposed in the intertemporal bar. Generally, however, postorbital-parietal overlap in Tropidostoma, when present, occurs throughout the length of the intertemporal bar (Fig. 7A) instead of in a ‘pinched’ posterior span. The posterior ramus of the postorbital terminates just beyond the point of the occipital plate (Fig. 4B).
The preparietal is a small, median element anterior to the pineal foramen (Fig. 4B). It is weakly depressed relative to the surrounding bone, taking into account that the entire surrounding region is strongly depressed (as discussed in the description of the frontal above). Preparietal depression is common in cryptodonts, and is also present in Tropidostoma. However, in Tropidostoma this bone is typically a narrow, anteroposteriorly elongate element with a pointed anterior tip (Fig. 7A). In Bulbasaurus, the preparietal is relatively wide, with a broadly rounded anterior tip (Fig. 4B), as in Aulacephalodon and Pelanomodon. The pineal foramen is a small, subcircular opening situated between the parietals (posteriorly and laterally) and preparietal (anteriorly). It is not elevated on a mound-like boss (as in rhachiocephalids, Endothiodon, and some large specimens of Aulacephalodon), but does have a raised, collar-like rim (Fig. 8). As noted above, the parietals are largely obscured by the postorbitals within the intertemporal bar in several specimens of Bulbasaurus. When exposed, their surface is weakly concave and otherwise unornamented.
As is usual in dicynodonts, the vomer is exposed only within the interpterygoid vacuity in ventral view (Fig. 5B). Anteriorly it is a narrow, rod-like element confluent with the median palatal ridge of the premaxilla. Posteriorly is slopes dorsally and bifurcates, developing a median trough before terminating in paired, flattened rami pressed against the medial walls of the anterior pterygoid rami.
The palatine is exposed ventrally in the form of a palatine pad, a roughly teardrop-shaped structure that would have formed part of the masticatory surface of the palate, and a shelf-like portion posterolateral to the pad that braces the medial edge of the anterior pterygoid ramus (Fig. 5B). The transversely expanded portion of the palatine pad is highly rugose, indicative of keratinous covering. Anterior to this is a short, sloping stretch of palatine contacting the premaxilla, with smooth bone texture. The palatine shelf attenuates posteriorly, extending along the medial wall of the pterygoid; this portion of the bone also has smooth bone texture. Although attenuate, this posterior portion is relatively thick in Bulbasaurus (Figs. 5B, 7E) compared to that of Tropidostoma (Fig. 7D) or Oudenodon, but is comparable to that of Aulacephalodon (Fig. 7F). In addition to its palatal contribution, the palatine is also exposed posterolaterally, behind the jugal and above the anterior pterygoid ramus along the rim of the subtemporal fenestra. Here, it contributes to the posterior rim of the labial fossa.
The ectopterygoid is a small, laminar element bordering the lateral wall of the anterior pterygoid ramus (Figs. 3B and 5B). It curves slightly laterally anteriorly, where it forms part of the border of the labial fossa. The anterior ramus of the pterygoid is a robust structure in Bulbasaurus, as in other geikiids. In Tropidostoma, the anterior pterygoid ramus is relatively narrower, with weaker lateral splay, and is generally more elongate (Fig. 7D). Thin crests on the ventral edges of the anterior pterygoid rami converge into the crista oesophagea posteriorly, on the median pterygoid plate (Fig. 9). The crista oesophagea is very well developed in Bulbasaurus, forming a tall, blade-like structure. Finally, the quadrate ramus of the pterygoid is a rod-like process, highly splayed outwards (57° relative to the long axis of the skull in the holotype) to contact the quadrate posterolaterally.
Sutures in the braincase of Bulbasaurus are not evident in any of the known specimens; it is likely that the braincase elements are extensively fused. Posterior to the median pterygoid plate, diverging ridges on what should be the parabasisphenoid (by comparison with other dicynodonts; King, 1988) extend towards the basal tubera (Fig. 5B). Each tuber is a thick, crescentic structure surrounding a ventrolateral exposure of the fenestra ovalis. The stapes is preserved in the specimen CGP/1/2263 (Fig. 15D). It is a stout, dumbbell-shaped element similar to those of other cryptodonts. A deep depression separates the basal tubera, and no intertuberal ridge is present. Posterior to this depression is the occipital condyle, which has the typical tripartite morphology of dicynodonts (presumably composed of a ventral basioccipital portion and paired dorsal exoccipital portions, although these elements are completely fused in Bulbasaurus). Circular jugular foramina are present ventrolateral to the occipital condyle, as is also typical of dicynodonts (Fig. 5B). Dorsal to the occipital condyle, the foramen magnum is ovoid, taller than wide (Figs. 6B and 11).
The occiput of SAM-PK-K11235 is well-prepared and fairly well-preserved (other than the broken edges of the squamosals), and clearly shows that most of the occipital elements are fused (Fig. 6B). Fusion between the opisthotic and prootic to form a periotic bone is common in dicynodonts, and incorporation of additional braincase bones is also observed in many taxa (Kammerer, Angielczyk & Fröbisch, 2015a). In Bulbasaurus, the occipital/braincase element is formed from fusion of the supraoccipital, exoccipitals, basioccipital, opisthotic, and prootic. The supraoccipital portion of this fused element is unusually expansive, making up the majority of the non-squamosal area of the occiput above the foramen magnum. The supraoccipital is typically a large element in dicynodonts, but usually the postparietal is nearly equal in size and the occiput has substantial contributions by the tabulars (this is the condition in Tropidostoma, for instance). In Bulbasaurus, the postparietal and tabulars are substantially smaller than the supraoccipital. The tabular is an arcuate, paired element between the squamosal, supraoccipital, and postparietal. Unusually for a therapsid, its long axis is horizontal rather than vertical. The most unusual occipital element is the postparietal, a median element at the top of the occipital plate. In most dicynodonts the postparietal is a flat, laminar element in-plane with the rest of the occiput, although it may have a dorsal process extending onto the skull roof (prominently in oudenodontids and especially so in cistecephalids). In Bulbasaurus, however, the entire postparietal is out-of-plane with both the skull roof and occipital plate, instead forming a sharply-angled ‘divot’ in the back of the skull. A very strongly developed nuchal crest is present along the postparietal midline and restricted entirely to that element.
The mandible is only preserved in two specimens of B. phylloxyron: CGP/1/970 (Figs. 12–14) and SAM-PK-K10106, and is largely unprepared in the latter. In CGP/1/970, the mandible is partially occluded to the cranium, but is somewhat dislocated (Fig. 14), revealing details of its dorsal surface (Fig. 13). In general, the mandible is similar to that of Aulacephalodon. The dentaries are fused to form an edentulous beak. The anterior face of the jaw symphysis is smoothly convex, without an anterior median ridge (Fig. 14). A sharp ridge delimits the edge between the anterior face of the symphysis and the lateral face of the jaw ramus (Fig. 12B), which is also present in Pelanomodon and Geikia but not Aulacephalodon. The anterodorsal terminus of the jaw comes to a sharp, pointed tip with a concave posterior surface. Although lower in dorsoventral height than the symphysis, the lateral portion of the jaw ramus is still tall and robust. The mandibular fenestra is a narrow, oval opening at mid-height on the dentary ramus; it is directly overhung by a small lateral dentary shelf that expands anterodorsally to form a broadened, diffuse boss. Unfortunately, the postdentary bones are not well-preserved in CGP/1/970; what is present is indistinguishable from that of other geikiids. Ventrally, the right articular is partially exposed, and has a well-developed, bulbous retroarticular process (Fig. 14).
The postcranial elements preserved in CGP/1/970 consist of a partially disarticulated set of cervical vertebrae, ribs, pectoral girdle, and forelimb elements (Figs. 13 and 14). Of these elements only the ribs (Fig. 13) and a humerus (Fig. 14) are reasonably exposed. The proximal ends of two left and four right ribs are preserved. They are bicipital and gently curved along the shaft, as in other dicynodonts. The humerus appears to represent a right humerus (visible as the large element at the bottom of Fig. 14). Its distal tip is broken off but it is otherwise intact, albeit extensively obscured by matrix and other bones. The deltopectoral crest is robust and strongly separated from the humeral shaft at a perpendicular angle.
Discussion
Ontogeny of Bulbasaurus
Most specimens of Bulbasaurus phylloxyron occupy the size range of 13–16 cm dorsal skull length. Two specimens fall outside of this range: CGP/1/2263 (Fig. 15) at 10.9 cm and SAM-PK-K10587 (Fig. 16) at 10.4 cm dorsal skull length. The short skull of CGP/1/2263 is at least partially artifactual, as this specimen is strongly anteroposteriorly compressed. SAM-PK-K10587, however, represents a genuinely smaller skull (in all dimensions) than other specimens of Bulbasaurus. Intriguingly, this specimen also differs from the larger specimens in a suite of cranial features, providing insight into the possible ontogenetic trajectory of Bulbasaurus.
Although the snout tip of SAM-PK-K10587 is still noticeably deflected (Figs. 16B and 16E), the snout in general is shorter and less ‘hook’-like than in the larger specimens of Bulbasaurus. The tusks of SAM-PK-K10587 are large and already-erupted (Fig. 16D), unlike Aulacephalodon or Tropidostoma skulls of equivalent size, but are on the low end of tusk proportions for Bulbasaurus (Table 2). The nasal bosses are weakly developed (Fig. 16A) and more clearly separated by a wider intervening ascending process of the premaxilla (Fig. 16F) than in the larger specimens. The interorbital region is proportionally narrower than in larger specimens, whereas the intertemporal region is comparatively broader (Fig. 16A). Although the curvature of the intertemporal region is already evocative of having a posterior ‘pinched’ portion in SAM-PK-K10587, no overlap of the postorbital on the parietal is present. There is also relatively little depression of the interorbital and intertemporal region, with the preparietal even being somewhat raised relative to the surrounding flat frontoparietal region (Fig. 16A). Finally, the zygomatic ramus of the squamosal is only slightly deflected at its posterodorsal edge; the subtemporal bar is not twisted 90°(Figs. 16B and 16E).
Most of the differences between SAM-PK-K10587 and the larger Bulbasaurus skulls are also observed in the ontogeny of Aulacephalodon (Tollman, Grine & Hahn, 1980; Kammerer, Angielczyk & Fröbisch, 2015a). The main exception is in the width of the intertemporal bar, which seems to follow the ancestral cryptodont pattern in Bulbasaurus (increasing overlap of the parietals by the postorbitals with growth, as is also observed in oudenodontids and rhachiocephalids) rather than the geikiid pattern (expansion of intertemporal parietal width with growth; Kammerer, Angielczyk & Fröbisch, 2015a). Remarkably, development of the complete suite of ‘adult’ geikiid features in Bulbasaurus occurs between skulls differing only in ∼3 cm length, whereas in Aulacephalodon this degree of ontogenetic change is observed between skulls differing in 10 cm length or more. Presence of these features in ∼13 cm long cranial specimens of Bulbasaurus suggests that most ‘adult’ characteristics of geikiids were evolved at small size and retained by giant descendants, not evolved in concert with the adoption of large body size.
Comparisons with other nominal cryptodont species
Twentieth century Karoo therapsid workers, especially Robert Broom, established a superfluity of dicynodont taxon names, the majority of which have subsequently been synonymized (King, 1988; Wyllie, 2003; Kammerer, Angielczyk & Fröbisch, 2011). Most of these synonymies have proven uncontroversial, particularly in the cases of supposed sympatric ‘species’ whose holotypes are distinguished solely by degree of taphonomic distortion. However, recent studies of dicynodont anatomy and stratigraphic distribution have revealed that the latter-day spate of taxonomic lumping may have been a bit overzealous, and that some previously-synonymized taxa are better recognized as distinct morphospecies (e.g., Angielczyk, 2002; Kammerer, Angielczyk & Fröbisch, 2015b). Given the vast array of available names for Karoo dicynodonts, it is important to consider whether the novel morphotype recognized herein could belong to a preexisting nominal species. Comparisons with well-known, valid species such as Tropidostoma dubium and Aulacephalodon bainii have already been presented in the context of the Description, but in this section, additional attention will be given to nominal species currently lurking in synonymy lists and species inquirenda.

Figure 17: Holotypes of nominal species currently considered synonymous with Tropidostoma dubium.
TM 249, holotype of Cteniosaurus platyceps Broom, 1935, in (A) dorsal and (B) right lateral view. TM 252, holotype of Dicynodon validus Broom, 1935, in (C) left lateral view. SAM-PK-2356, holotype of Dicynodon rogersi Broom & Haughton, 1917, in (E) dorsal and (D) right lateral view. TM 250, holotype of Dicynodon acutirostris Broom, 1935, in (F) left lateral view. SAM-PK-747, holotype of Dicynodon cavifrons Broom & Haughton, 1917, in (H) dorsal and (G) right lateral view. NHMUK 47051, holotype of Dicynodon dubius Owen, 1876, in (I) dorsal and (J) left lateral view. Scale bars equal 5 cm.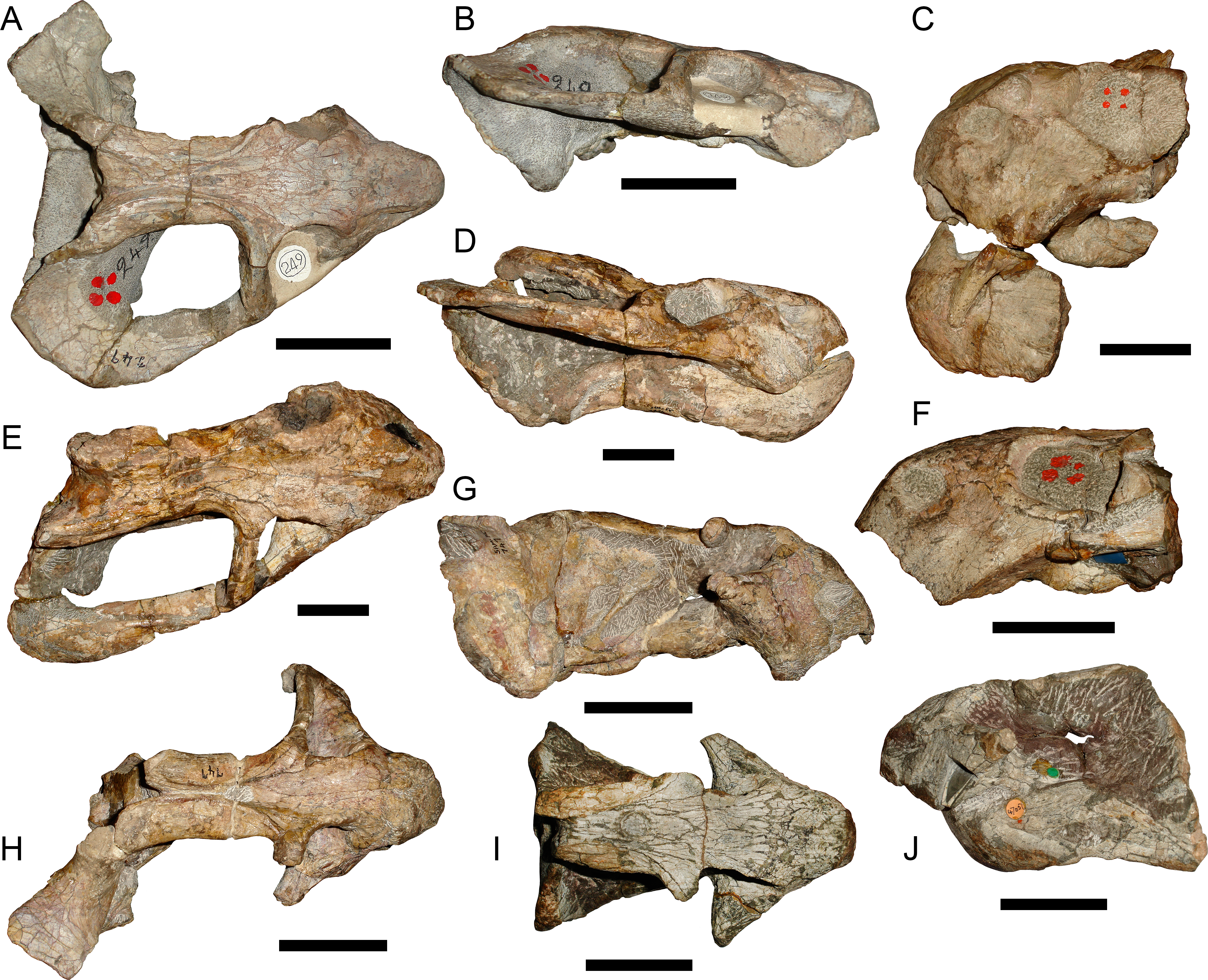{kind=link}
Prior to the recognition of Bulbasaurus phylloxyron as a distinct morphotype, all of the specimens of this taxon were identified in collections as Tropidostoma. As such, the most likely source for potential preexisting names representing this morphotype is among the nominal species currently considered synonymous with Tropidostoma dubium (Keyser, 1973; Botha & Angielczyk, 2007; Kammerer, Angielczyk & Fröbisch, 2011). Seven nominal dicynodont species are currently in synonymy with T. dubium: Tropidostoma dunnii Seeley, 1889 (the type species of Tropidostoma), Dicynodon microtrema Seeley, 1889, Dicynodon cavifrons Broom & Haughton, 1917, Dicynodon rogersi Broom & Haughton, 1917, Cteniosaurus platyceps Broom, 1935, Dicynodon acutirostris Broom, 1935, and Dicynodon validus Broom, 1935. Within the current hypodigm for Tropidostoma dubium, two cranial morphs can be recognized: one ‘robust’ with a relatively tall, short snout and large tusks, and one ‘gracile’ with a relatively long, low snout and small tusks. The holotypes of Dicynodon dubius (Figs. 17I, 17J), D. cavifrons (Figs. 17G, 17H), and D. validus (Fig. 17C) correspond to the ‘robust’ morph, whereas the holotypes of C. platyceps (Figs. 17A, 17B), D. rogersi (Figs. 17D, 17E), and D. acutirostris (Fig. 17F) correspond to the ‘gracile’ morph. It is possible that this morphological disjunct reflects taxonomic distinction, but given that similar morphs are seen in the confamiliar Oudenodon bainii (C Kammerer, pers. obs., 2016) it is more likely that this represents sexual dimorphism, a not-uncommon occurrence in dicynodonts (Tollman, Grine & Hahn, 1980; Sullivan, Reisz & Smith, 2003; Ray, 2005; Kammerer, Angielczyk & Fröbisch, 2015a; Kammerer, Angielczyk & Fröbisch, 2015b). Whether these morphs exhibit the frequency expected for sexual dimorphism and whether they are truly dimorphs or represent a cline of individual variation will be addressed in a future contribution. Importantly for the topic at hand, however, neither of these morphs corresponds to the morphology of Bulbasaurus.
The holotypes of T. dunnii (NHMUK R866) and D. microtrema (NHMUK R868) consist solely of isolated occipital plates with attached intertemporal bars. Although the incompleteness of these specimens limits comparisons, they can clearly be differentiated from Bulbasaurus by their intertemporal morphology: both specimens have elongate intertemporal bars in which the postorbitals almost completely overlap the parietals for the extent of their length (similar to that of Fig. 7A). By contrast, in Bulbasaurus postorbital overlap of the parietals is limited to a posterior ‘pinched’ portion of the intertemporal bar (Fig. 4) if it is present at all. The holotype of D. cavifrons (SAM-PK-747) is a well-preserved partial skull missing all of the the left and most of the right temporal arches. This specimen is roughly similar to Bulbasaurus in having relatively large tusks and a deep snout (Fig. 17G), characters shared with other ‘robust’ morph Tropidostoma specimens. However, it can be distinguished from Bulbasaurus by the elongate intertemporal bar, with almost complete postorbital-parietal overlap along its length, the presence of large, triangular postfrontals, a relatively narrow interorbital region, small, ovoid nasal bosses restricted to overhanging the external nares (Fig. 17H), the presence of postcaniniform crests, and thin quadrate rami of the pterygoids. The same features also distinguish the holotypes of D. rogersi (SAM-PK-2356, a dorsoventrally compressed partial skull missing the left temporal arch) and C. platyceps (TM 249, a partial skull missing most of the left temporal arch) from Bulbasaurus, with the exception of there being a broad intertemporal exposure of the parietal in these specimens (Figs. 17A and 17E). These specimens can be further differentiated from Bulbasaurus by their transversely narrow snouts (Figs. 17A and 17E) and relatively small tusks. The holotype of D. acutirostris (TM 250, an isolated snout) also has a transversely narrow snout, but further differs from Bulbasaurus in the complete absence of tusks (Fig. 17F). The holotype of D. validus (TM 252, a partial snout with the anterior portion of the lower jaws) is another ‘robust’ morph skull, but can readily be distinguished from Bulbasaurus by nasal boss morphology, with a highly discrete ovoid boss restricted to the posterodorsal edge of the external naris (Fig. 17C). Finally, it is worth making comparisons to the holotype of Tropidostoma dubium (formerly Dicynodon dubius) itself, as the contrasts with T. dubium in the Description were based on exemplary referred specimens rather than the type. The holotype (NHMUK 47051) is a partial skull and occluded lower jaws, missing the temporal arches. Very large, triangular postfrontals are present in this specimen, and the nasal bosses exhibit the morphology typical for Tropidostoma (Figs. 17I and 17J). Furthermore, in this specimen the tusks were just beginning to erupt at the time of death (visible within the ground-off caniniform process; see Fig. 17J), despite being a larger individual (dorsal skull length: 11.1 cm) than SAM-PK-K10587, which although the smallest (dorsal skull length: 10.4 cm) specimen of Bulbasaurus already has large, fully-erupted tusks (Fig. 16).
Few specimens comparable to Bulbasaurus are present among the type series of other synonym-rich cryptodonts. The many synonyms of Oudenodon bainii can be distinguished from Bulbasaurus by their consistent lack of tusks, in addition to various shared oudenodontid characters that also distinguish Tropidostoma from Bulbasaurus (such as general gracility of the pterygoids and intertemporal morphology). Almost all holotypes referable to Aulacephalodon bainii are large specimens exhibiting the full suite of features that distinguish adults of that species from Bulbasaurus (e.g., extremely broad intertemporal region, massive but discrete nasal bosses protruding over the external nares, bosses or swellings on the zygomatic arch). A notable exception, however, is BP/1/763, a dorsoventrally compressed complete skull that forms the holotype of Proaulacocephalodon miltoni Toerien, 1955. Tollman, Grine & Hahn (1980) considered BP/1/763 to represent a juvenile individual of Aulacephalodon bainii, and attributed its very small tusks, narrow intertemporal and interorbital regions, and lack of nasal and prefrontal bosses to an early stage of ontogeny. This specimen is indeed relatively small (dorsal skull length: 15.6 cm) compared with most skulls of A. bainii. However, the recent discovery of even smaller specimens (notably SAM-PK-K11409, dorsal skull length: 10.2 cm; formerly B 168, see Kammerer, Angielczyk & Fröbisch, 2015a) referable to A. bainii that already show distinct, protruding nasal bosses and intertemporal morphology similar to that of adult Aulacephalodon suggests that these features cannot be explained by ontogeny in BP/1/763. The proper attribution of BP/1/763 is uncertain at present, but it is probably not referable to Aulacephalodon. It is also not conspecific with Bulbasaurus phylloxyron, being distinguished by the proportionally shorter intertemporal region, narrower interorbital region, absence of nasal bosses, extremely small tusks (despite being an absolutely larger skull than nearly all specimens of Bulbasaurus), and relatively thin quadrate rami of the pterygoids. A similarly problematic specimen is TM 1480, holotype of Dicynodon hartzenbergi Broom, 1940. This specimen consists of a skull and lower jaws, with the anterior tip of the snout worn off. Brink (1986) considered TM 1480 to be a juvenile of Aulacephalodon, but the absence of protruding nasal bosses and tusks makes this identification unlikely. Kammerer, Angielczyk & Fröbisch (2011) noted some similarities with geikiids, but left this taxon as a species of uncertain attribution. In overall skull proportions this specimen is similar to that of Bulbasaurus, but differs from the latter taxon in the absence of tusks, substantially narrower interorbital region, and apparent restriction of the small nasal bosses to the edges of the external nares. In these characters it is similar to Oudenodon, and this specimen may eventually prove to be an anteroposteriorly-deformed specimen of O. bainii, although further research is needed. In addition to the morphological features listed above, from a stratigraphic standpoint it is unlikely that either Proaulacocephalodon miltoni or Dicynodon hartzenbergi are conspecific with Bulbasaurus phylloxyron, because neither are from the Tropidostoma AZ, both hailing from the overlying Cistecephalus AZ (with BP/1/763 found in Windpoort, Murraysburg, Western Cape Province, and TM 1480 found in Petersburg, Graaff-Reinet, Eastern Cape Province).
Relationships of Bulbasaurus
Bulbasaurus phylloxyron was included in a modified version of the anomodont phylogenetic analysis of Kammerer, Angielczyk & Fröbisch (2011), incorporating data from the revisions of Castanhinha et al. (2013), Kammerer, Angielczyk & Fröbisch (2013), Kammerer, Angielczyk & Fröbisch (2015b), Kammerer, Bandyopadhyay & Ray (2016), Angielczyk et al. (2016), and Boos et al. (2016). This analysis includes 21 continuous characters, 153 discrete state characters, and 103 operational taxonomic units. Codings for Bulbasaurus were based on all known specimens, with variation between specimens coded as polymorphism and continuous characters coded as means (see Data S1). Analyses were run in TNT v1.1 (Goloboff, Farris & Nixon, 2008) using New Technology search parameters (sectorial search, parsimony ratchet, drift, and tree fusing) with an initial driven search level of 65 and the requirement of finding the most parsimonious tree 20 times. Bootstrap support was calculated based on 10,000 replicates.
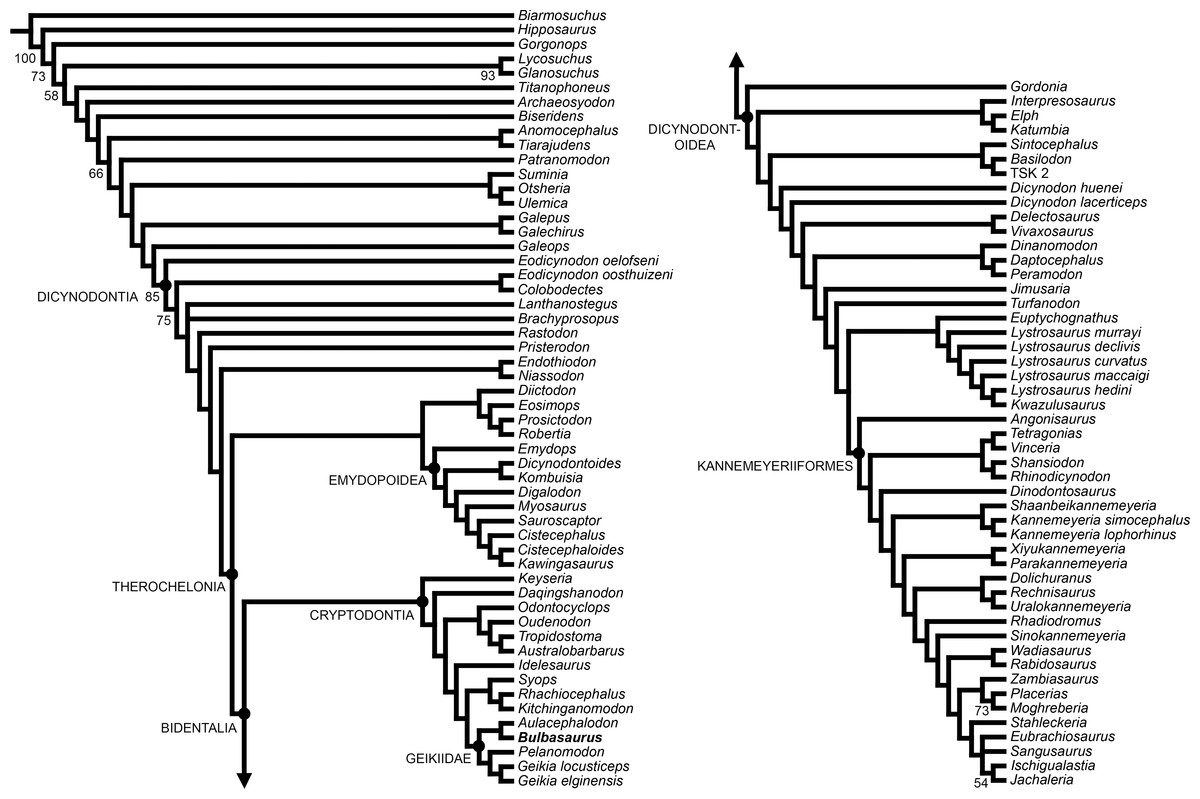
Figure 18: Strict consensus of the two most parsimonious trees resulting from the phylogenetic analysis.
Major clades discussed in the text are labeled at their respective nodes; numbers at nodes represent bootstrap values >50.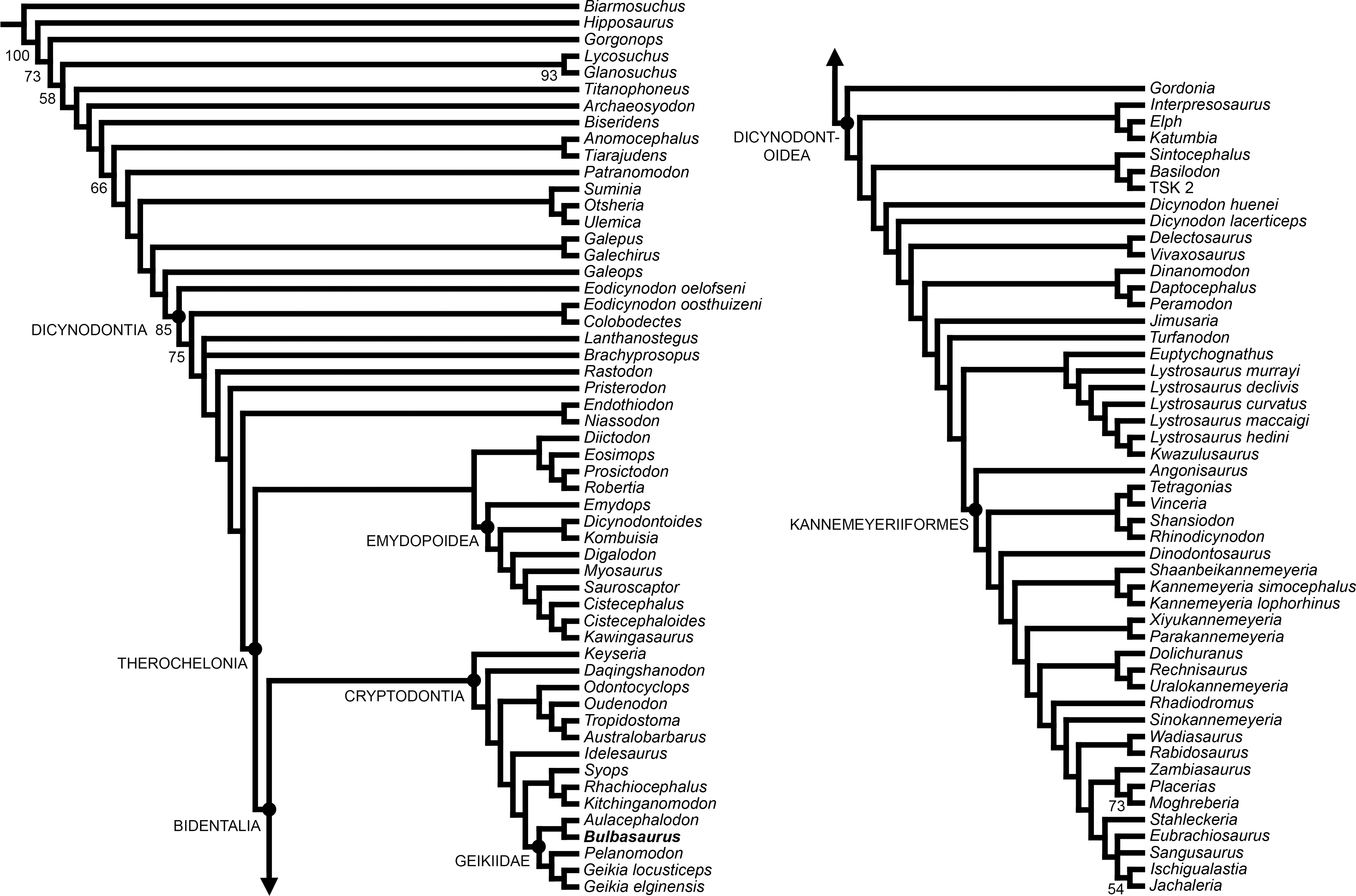{kind=link}
Two most parsimonious trees of length 1032.706 were recovered, with a consistency index of 0.234 and a retention index of 0.709 (Fig. 18). The only differences between the two trees are in the positions of the middle Permian taxa Brachyprosopus broomi and Lanthanostegus mohoii (which swap positions as successive outgroups to the clade containing Rastodon, Pristerodon, Endothiodontia, and Therochelonia) and the Triassic stahleckeriids Eubrachiosaurus and Sangusaurus (which swap positions as the sister-taxon of Ischigualastia + Jachaleria). The current tree differs from that of the recent analysis by Boos et al. (2016) in several important regards. Boos et al. (2016) recovered the newly-described middle Permian Brazilian dicynodont Rastodon as the earliest-diverging member of Bidentalia. In the current analysis Rastodon is recovered in a far more rootward position, as the sister-taxon to all dicynodonts other than Eodicynodon, Colobodectes, Brachyprosopus, and Lanthanostegus. Pylaecephalidae (the family containing Diictodon and its close relatives) is recovered within Therochelonia, as the sister-taxon to Emydopoidea, as in the majority of recent analyses (Kammerer, Angielczyk & Fröbisch, 2011; Kammerer, Angielczyk & Fröbisch, 2013; Kammerer, Angielczyk & Fröbisch, 2015b; Kammerer, Bandyopadhyay & Ray, 2016; Castanhinha et al., 2013; Cox & Angielczyk, 2014; Angielczyk et al., 2016). By contrast, Boos et al. (2016) recovered Pylaecephalidae outside of Therochelonia, as in a number of earlier analyses (Angielczyk, 2002; Angielczyk, 2007; Angielczyk & Kurkin, 2003; Fröbisch, 2007; Fröbisch & Reisz, 2008; Angielczyk & Rubidge, 2013).
Cryptodontia in its traditional sense (Kammerer & Angielczyk, 2009) was recovered as monophyletic (contra other recent iterations of this analysis; e.g., Boos et al., 2016; Kammerer, Bandyopadhyay & Ray, 2016). Keyseria and Daqingshanodon were recovered as the earliest-diverging cryptodonts, as in all other analyses in which Cryptodontia is monophyletic (e.g., Kammerer, Angielczyk & Fröbisch, 2011; Kammerer, Angielczyk & Fröbisch, 2013; Castanhinha et al., 2013; Cox & Angielczyk, 2014). The three recognized cryptodont families (Oudenodontidae, Rhachiocephalidae, and Geikiidae) were each recovered containing their ‘core’ taxa (i.e., Oudenodon, Australobarbarus, and Tropidostoma in Oudenodontidae; Rhachiocephalus and Kitchinganomodon in Rhachiocephalidae; and Geikia, Aulacephalodon, and Pelanomodon in Geikiidae). Unusually, however, Odontocyclops was recovered as an oudenodontid and Syops was recovered as a rhachiocephalid. Both of these results are novel: Odontocyclops was previously recovered as either the sister-taxon of Idelesaurus in a clade outside (Geikiidae + Rhachiocephalidae) (Kammerer, Angielczyk & Fröbisch, 2011; Kammerer, Angielczyk & Fröbisch, 2013; Castanhinha et al., 2013; Cox & Angielczyk, 2014) or as a geikiid (Boos et al., 2016) and Syops was previously recovered as either a dicynodontoid (Kammerer, Angielczyk & Fröbisch, 2011; Boos et al., 2016) or a geikiid (Castanhinha et al., 2013; Kammerer, Angielczyk & Fröbisch, 2013; Cox & Angielczyk, 2014). The position of Odontocyclops is based on five continuous characters (character 8, length of interpterygoid vacuity; character 12, ratio of length to height of mandibular fenestra; character 15, relative minimum and maximum widths of scapula; character 17, maximum width of distal end of radius; and character 18, ratio of posterior iliac process length to acetabular diameter) and two discrete state characters (character 52, state 1: small postparietal contribution to the skull roof, present in the other oudenodontids as well as Kitchinganomodon; and character 106, state 0: absence of ridge on edge of dentary symphysis, also coded for the other oudenodontids, Syops, Rhachiocephalus, Kitchinganomodon, and Aulacephalodon). The position of Syops is based on one continuous character (character 4: relative width of intertemporal bar) and two discrete state characters (character 46, state 2: parietal exposed in intertemporal bar as a thin crest, otherwise only coded for Rhachiocephalus and Kitchinganomodon among cryptodonts; and character 106, state 0: absence of ridge on edge of dentary symphysis).
Bulbasaurus was recovered as a geikiid in this analysis, sister-taxon to Aulacephalodon. The position of Bulbasaurus as a geikiid is supported by four discrete state characters (character 11, state 1: marked anterior expansion of preorbital region; character 34, state 1: transverse nasofrontal ridge; character 39, state 1: absence of postfrontal; and character 51, state 2: pineal foramen surrounded by raised edge). Its position as sister-taxon to Aulacephalodon is based solely on the absence of a postcaniniform crest (character 29, state 0). A postcaniniform crest on the posteroventral surface of the caniniform process is present in all other known cryptodonts. However, as discussed above, the postcaniniform crest is also present early in ontogeny in Aulacephalodon and lost when the tusks expand to full size, causing the caniniform process to bulge outwards. If the tusks did not develop in an Aulacephalodon-like animal, one would expect the postcaniniform crest to be retained. As such, this synapomorphy is questionable, considering the absence of tusks in Pelanomodon and Geikia. Indeed, it requires only a single additional step for Bulbasaurus to fall outside of Geikiinae (the clade containing Aulacephalodon, Geikia, and Pelanomodon) as a basal geikiid, a position better reflecting stratigraphy. Additional research on the patterns of character evolution in Cryptodontia is required to test this possibility. Cryptodonts in general suffer from a high degree of topological instability, low character support, and rampant homoplasy (as reflected in low consistency indices and extremely variable positions for taxa such as Syops), indicating that the current phylogenetic data set is not sufficient to robustly resolve their relationships.
Conclusions
Bulbasaurus phylloxyron is identified as a new taxon of dicynodont, representing the earliest known record for the family Geikiidae and eliminating the ghost lineage for this clade in the Tropidostoma AZ. Bulbasaurus is most similar among known dicynodonts to its later relative Aulacephalodon, and exhibits many cranial characteristics of the latter taxon at substantially smaller skull size (13–16 cm dorsal skull length). The presence of ‘adult’ Aulacephalodon features in relatively small skulls suggests that these specimens represent mature individuals of Bulbasaurus, and that geikiids started out as relatively small animals. Similarity in size to Tropidostoma and absence of some of the more obvious features of later geikiids (e.g., broadened intertemporal region, protruding nasal bosses) previously obscured the true identity of Bulbasaurus specimens in collections. With the recognition of this taxon, however, Geikiidae can be added to the list of bidentalians (with Oudenodontidae and Rhachiocephalidae already represented) that first appear as rare faunal components of the Tropidostoma AZ. The continuing absence of dicynodontoids in this fauna remains problematic. Additional research attention should be given to revising existing collections of Tropidostoma AZ dicynodonts, as it is possible that early dicynodontoids could also be hiding among these specimens.