
A new species of alien terrestrial planarian in Spain: Caenoplana decolorata
- Published
- Accepted
- Received
- Academic Editor
- Jean-Lou Justine
- Subject Areas
- Biodiversity, Molecular Biology, Taxonomy, Zoology
- Keywords
- Molecular identification, Alien species, Invasive species, Land flatworm
- Copyright
- © 2020 Mateos et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2020. A new species of alien terrestrial planarian in Spain: Caenoplana decolorata. PeerJ 8:e10013 https://doi.org/10.7717/peerj.10013
Abstract
Terrestrial planarians found in a plant nursery in Spain in 2012 are described as a new species, Caenoplana decolorata. Dorsally they are mahogany brown with a cream median line. Ventrally they are pastel turquoise fading to brown laterally. Molecular data indicate that they are a member of the genus Caenoplana, but that they differ from other Caenoplana species found in Europe. One mature specimen has been partially sectioned, and the musculature and copulatory apparatus is described, confirming the generic placement but distinguishing the species from other members of the genus. It is probable that the species originates from Australia.
Introduction
Álvarez-Presas et al. (2014) recorded several terrestrial planarian species from Spain, some considered native to Europe, others introduced from other continents. Some species were identifiable on the basis of external features such as colour and shape, on anatomical characters and comparative molecular analysis. Molecular results suggested that further species were found but at the time they could not be certainly identified to species, though perhaps to genus. This paper describes specimens (Figs. 1A–1D) listed as ‘Caenoplana Ca2’ by Álvarez-Presas et al. (2014). Molecular data (Fig. 12 of Álvarez-Presas et al., 2014) indicate that these specimens are of the genus Caenoplana Moseley, 1877, but distinct from other Caenoplana species. One mature specimen has been partially sectioned, and the musculature and copulatory apparatus is described. It has the characters of the genus Caenoplana Moseley, 1877, as amended by Ogren & Kawakatsu (1991) and by Winsor (1991) but differs from other described species of that genus both in external characteristics and anatomy. Neither do the specimens resemble any species described only on external features such as shape and colour and currently placed in the genus Australopacifica Ogren & Kawakatsu, 1991, a collective genus containing species ‘not classifiable into the present taxonomic genera because of insufficient morphological information; geographical distribution largely in Australasia and Indo-Pacific Islands. A collective group for species inquirendae and nomina dubia’. It is described as Caenoplana decolorata sp. nov.

Figure 1: Caenoplana decolorata sp. nov.
(A–D) Photographs of live specimens, anterior to the right. (A) Dorsal view of specimen PT426 showing the ‘mahogany brown’ colour and ‘cream’ median line. Scale bar 10 mm. (B) A twisted specimen PT657-1 showing the ‘pastel turquoise’ ventral surface. Scale bar 10 mm. (C) Specimen PT657-1 and (D) specimen PT426, anterior end showing anterior ‘copper brown’ colour and the eyes (the two white lines in (D) are reflections from the lighting). Scale bars 4 mm. (E and F) Pots under which the specimens were found, in a greenhouse (E) and outdoors (F).{kind=link}
Methods
Sampling
Specimens were collected by E. Mateos from a plant nursery named “vivers casa Paraire” in Bordils municipality (Girona province, Spain, WGS84, position: 42.0348°N; 2.8986°E). All were collected by hand from beneath pots (Figs. 1E and 1F) that contained the plants on 12 January 2012 (five specimens: PT426, PT427, PT428, PT430, PT431) and 22 October 2012 (four specimens: PT655, PT657-1, 2 and 3) (Table 1).
| Species/morphotype | Locality | GenBank Code |
|---|---|---|
| Cox1 | ||
| Family Geoplanidae | ||
| Subfamily Rhynchodeminae | ||
| Tribe Caenoplanini | ||
| Artioposthia sp. | Australia | MN990642 |
| Arthurdendyus testaceus | – | MN990643 |
| Caenoplana coerulea | New Zealand | DQ665961 |
| Menorca (Spain) | JQ514564 | |
| Liverpool, UK | DQ666030 | |
| El Prat de Llobregat (Barcelona, Spain) | KJ659617 | |
| Vall de’n Bas (Girona, Spain) | KJ659618 | |
| KJ659619 | ||
| KJ659620 | ||
| KJ659622 | ||
| KJ659623 | ||
| KJ659624 | ||
| KJ659626 | ||
| Badalona (Barcelona, Spain) | KJ659633 | |
| KJ659634 | ||
| Adelaide (Australia) | KJ659642 | |
| – | KJ659645 | |
| Granollers (Barcelona, Spain) | KJ659647 | |
| PT1304 | Real Jardín Botánico de Córdoba (Córdoba, Spain) | MT727076* |
| PT1305 | MT727077* | |
| PT1307 | MT727078* | |
| PT1310 | MT727079* | |
| Caenoplana sp. 1 | – | DQ666031 |
| Caenoplana sp. 2 | Tallaganda (Australia) | DQ227621 |
| DQ227625 | ||
| DQ227627 | ||
| DQ227634 | ||
| Caenoplana sp. 3 | Victoria (Australia) | DQ465372 |
| Caenoplana sp. 4 | – | DQ666032 |
| Caenoplana variegata | Bordils (Girona, Spain) | KJ659648 |
| Southampton, UK | MN990646 | |
| Cardiff, UK | MN990647 | |
| MN990648 | ||
| Caenoplana decolorata sp. nov. | Bordils (Girona, Spain) | KJ659628 |
| KJ659629 | ||
| KJ659630 | ||
| KJ659631 | ||
| KJ659632 | ||
| MN990644 | ||
| KJ659649 | ||
| OUTGROUP: tribe Rhynchodemini | ||
| Dolichoplana sp. | – | DQ666037 |
| D. striata | Igreginha (Brazil) | KC608226 |
| Rhynchodemus sylvaticus | Canyamars (Barcelona, Spain) | FJ969946 |
Note:
Specimens from 12 January 2012 and specimens PT655 and PT657-3 were preserved in absolute ethanol for further molecular analyses. Specimens PT657-1 and 2 were killed with boiling water, fixed with 10% formalin and preserved in 70% ethanol. Specimens PT426 and PT657-1 were photographed alive (Fig. 1).
Molecular methods
All the sequences used in the present work were obtained in previous studies with the exception of some Cytochrome Oxidase I (herein Cox1) sequences that were obtained from individuals collected at the Real Jardín Botánico de Córdoba (Spain) by Mónica López (Table 1). In all cases, a small section of the anterior end of specimens preserved in absolute ethanol was used for DNA extraction. The new sequences were obtained following the same protocol as in Álvarez-Presas et al. (2014).
A nucleotide alignment was obtained for the Cox1 sequences based on the AA translation as a guide using BioEdit software (Hall, 1999) and the echinoderm mitochondrial genetic code (9). A Maximum Likelihood (ML) phylogeny was inferred using the IQtree software (Nguyen et al., 2015) with the MFP+MERGE implementation and 10,000 replicates for ultrafast bootstrap search (-bb option). Then two single locus molecular species delimitation methods were applied in order to check the validity of the new species presented here and the ones already described and included in the phylogeny. Automatic Barcode Gap Discovery (ABGD) (Puillandre et al., 2012) was the first method performed, implemented in the webpage: https://bioinfo.mnhn.fr/abi/public/abgd/abgdweb.html. The default parameters were used, selecting initial partitions as they are supposed to be more stable and generally give as a result a closer number of groups described by taxonomists than recursive partitions. The second method applied was the multi-rate Poisson Tree Process (mPTP) analysis (Kapli et al., 2017). The newick tree obtained in the ML phylogenetic inference was used as input in the website http://mptp.h-its.org/#/tree.
Anatomical methods
Specimens PT657-1 and 2 were sent to HDJ and are deposited in the Natural History Museum, London, accession numbers NHMUK.2014.5.13.12-13. The larger specimen had a visible gonopore, was assumed to be mature and selected for partial sectioning. It was divided into four portions: anterior portion about 2 cm long not sectioned, in alcohol; pre-pharyngeal section, TS, five slides, two at 15 µm, three at 10 µm; posterior portion including pharynx and copulatory apparatus, LS, 16 slides (pharynx separated, HLS) at 15 µm. Sectioned portions were dehydrated in ethanol and embedded in paraffin wax. Slides were stained in Harris’s haematoxylin and eosin and mounted in Canada balsam. The second specimen, about 3.4 cm long, had no visible gonopore and remains in alcohol.
Colours are expressed as RAL colours (www.ralcolor.com).
The electronic version of this article in Portable Document Format (PDF) will represent a published work according to the International Commission on Zoological Nomenclature (ICZN), and hence the new names contained in the electronic version are effectively published under that Code from the electronic edition alone. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank Life Science Identifiers (LSIDs) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix http://zoobank.org/. The LSID for this publication is: urn:lsid:zoobank.org:pub:B2636DF8-4372-405C-8A8C-4FBEC7396276. The LSID for the new species described is: Caenoplana decolorata sp. nov. urn:lsid:zoobank.org:act:C0CEE92F-A51E-4EDD-B18B-E7F021338667. The online version of this work is archived and available from the following digital repositories: PeerJ, PubMed Central and CLOCKSS.
Results
Molecular results
The final dataset comprises 43 Cox1 sequences (including three outgroups, Table 1), with a final length of 822 bp. The resulting ML tree (Fig. 2) shows monophyletic groups comprising seven putative Caenoplana species. Although the bootstrap values (bb) are not high enough to give support to the relationships between these clades, the monophyly of the new species described here, C. decolorata, harbor maximum support. The results of the molecular species delimitation analyses (both mPTP and ABGD) match the same clades present in the phylogeny (Fig. 2) giving rise to seven putative Caenoplana species. Among them, we find the subject of this study, Caenoplana decolorata.

Figure 2: Maximum Likelihood (ML) phylogeny inferred with Cox1 sequences.
Values at nodes correspond to ultrafast bootstrap replicates (bb) obtained with IQtree software. Vertical bars to the right of the phylogeny correspond to the molecular species delimitation methods results (mPTP, left bar and ABGD, right bar). Scale bar represents number of substitutions per site. Photograph of specimen PT426 in dorsal view (anterior to the right).{kind=link}
Taxonomic section
Order Tricladida Lang, 1884
Suborder Continenticola Carranza et al., 1998
Family Geoplanidae Stimpson, 1857
Subfamily Rhynchodeminae von Graff, 1896
Tribe Caenoplaninae Ogren & Kawakatsu, 1991
Genus Caenoplana Moseley, 1877
Caenoplana decolorata new species.
Caenoplana Ca2 Álvarez-Presas et al., 2014.
Etymology: “decolorata” indicating that live specimens resemble C. coerulea but are comparatively pale and discolored.
NHMUK.2014.5.13.12-13
E. Mateos collection code PT657-1 and PT657-2. Locality: Bordils (Girona, Spain), position N42.0348049 E2.8986153, date 22 October 2012.
Preserved dimensions: holotype (PT657-1): length 46 mm; width 2 mm; height 1 mm; anterior to mouth 28 mm (61% of body length); anterior to gonopore 39 mm (85% of body length); paratype (PT657-2): length 34 mm; width 2.1 mm; anterior to mouth 17 mm (50%); apparently immature.
All other specimens (with a small section of the anterior end removed) are deposited in the collection of M. Riutort at the University of Barcelona.
External characters
Live specimens are ‘mahogany brown’ (RAL 8106) with a narrow ‘cream’ (RAL 9001) mid-line dorsally, merging to ‘beige brown’ (RAL 8024) laterally. The anterior end is ‘copper brown’ (RAL 8004). The ventral mid-line is ‘pastel turquoise’ (RAL 6034) merging into the lateral ‘beige brown.’
Eyes in a sparse uniserial row round the anterior end, biserial for a short distance behind the anterior end and sparse staggered uniserial to the posterior end. Sole nearly the whole of the ventral surface.
Anatomy
Transverse sections (Fig. 3A) are about 1.3 mm high and 2 mm wide. The ciliated creeping sole is about 80% of the width. The cilia are about 5 µm long. The ventral epidermis is a monolayer about 30 µm thick and has few rhabdites. Ventral sub-epidermal muscle consists of a layer of circular muscle fibres about 10 µm thick and longitudinal muscle in bundles about 30 µm thick. Dorsal to the longitudinal muscle bundles is a ventral nerve plexus. There is a distinct, compact layer of parenchymal longitudinal muscle ventrally, 40–50 µm thick, 150 µm in from ventral surface. Ventral nerve cords are about 750 µm centre to centre, about 120 µm in diameter, with transverse commissures. Laterally and dorsally the parenchymal longitudinal muscle is less compact and 10–20 µm thick. Dorsal epidermis is 45 µm thick, non-ciliated and has numerous rhabdites. Dorsal and lateral sub-epidermal circular muscle is about 10 µm thick, and longitudinal muscle in bundles about 35 µm thick. Rhabdites are numerous dorsally and laterally ental to the sub-epidermal muscle, but in the mid-dorsal region, the rhabdites layer is slightly deeper (Figs. 3A and 3B), presumed to be coincident with the pale midline visible in the living animal.
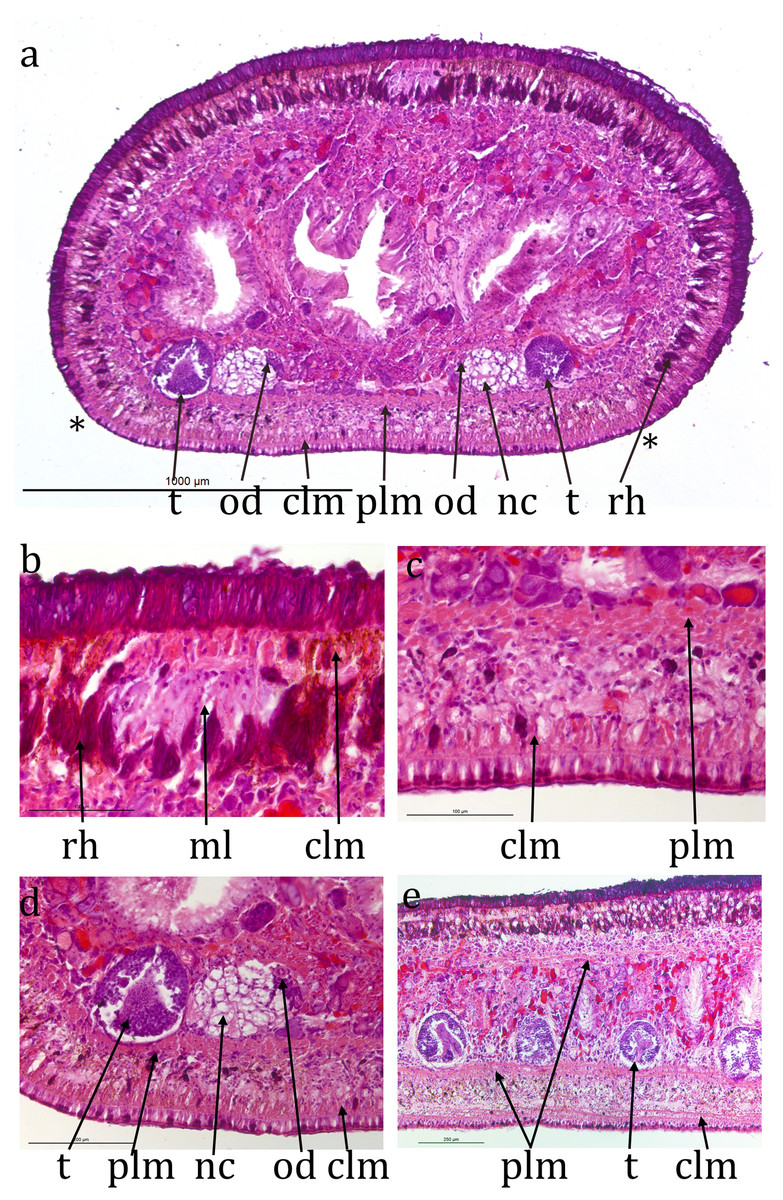
Figure 3: Caenoplana decolorata specimen PT657-1 (NHMUK2014.5.13.12).
(A) Entire transverse section (**indicate the width of the ventral creeping sole; scale line 1 mm). (B) Enlarged mid-dorsal (scale line 100 µm). (C) Enlarged mid-ventral (scale line 100 µm). (D) The testis, ventral nerve cord and ovovitelline duct on one side (scale line 200 µm). (E) Longitudinal section showing several testes (scale line 250 µm).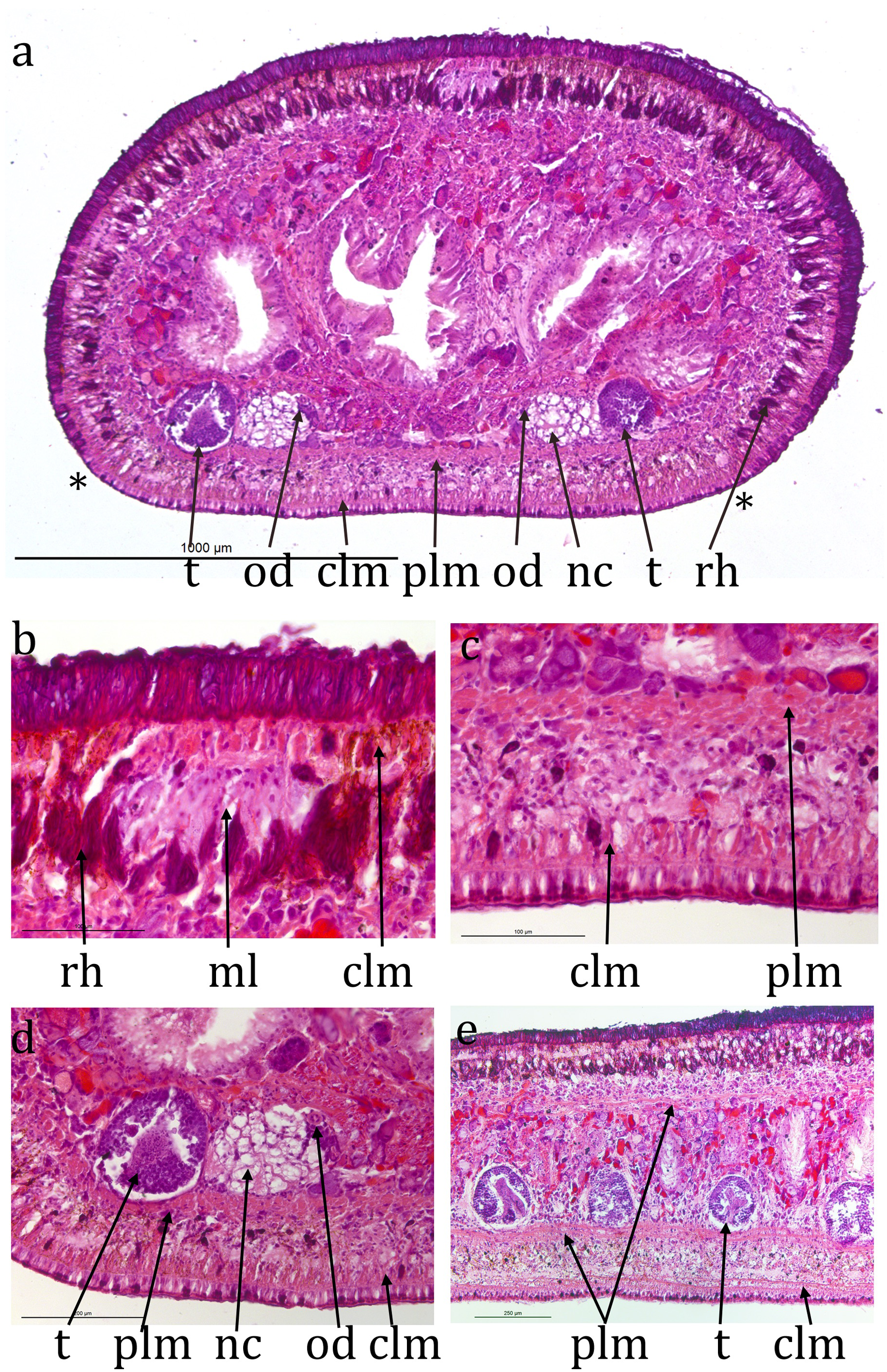{kind=link}
The retracted cylindrical pharynx occupies the whole length of the pharyngeal pouch and is about 2.5 mm long, 0.9 mm in diameter. The pouch is 5.4% of body length. The pharyngeal aperture is about half way along the pharyngeal pouch. Pharyngeal musculature consists of an outer layer of circular muscle about 10 µm thick, a layer of mixed longitudinal and radial muscle about 360 µm thick and an inner layer of circular muscle about 30 µm thick.
The anterior portion containing the ovaries has not been sectioned. Ovovitelline ducts are about 500 µm apart on the inner dorsal surface of the ventral nerve cords (Figs. 3A and 3D). Vitellaria are not distinguishable with certainty. Their outer and inner diameters are about 25 µm and 7 µm respectively. They run to about 800 µm behind the gonopore, turn dorsally and open into the common female duct about 800 µm long which extends forwards with little differentiation to open into the common antrum above the gonopore (Figs. 4A, 4C and 4E). There is little or no shell gland tissue surrounding the common female duct.
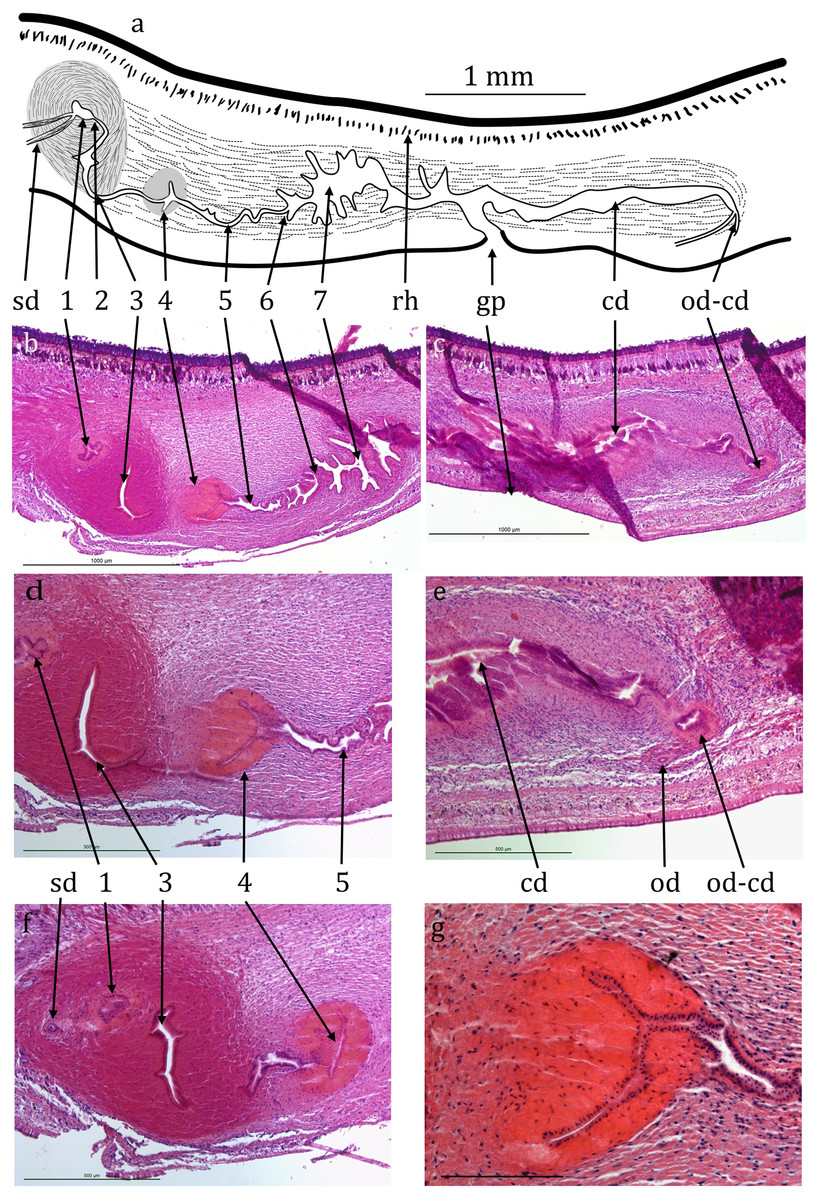
Figure 4: Caenoplana decolorata specimen PT657-1 (NHMUK2014.5.13.12).
(A) Reconstruction diagram and (D–G) longitudinal sections of the copulatory apparatus (anterior to the left); (A), (B) and (C) are to the same scale. Micrographs: (B and C) Mid-sections through the male and female portions respectively (both folded sections) (scale lines 1,000 µm). (D and F) Further sections through the proximal portion of the male ducts (scale lines 500 µm). (E) Section showing the approach of an ovovitelline duct to the common female duct (scale line 500 µm). (G) Enlargement of region 4 of the male duct (scale line 200 µm). The nuclei (cyanophil) are mostly adjacent to the lumen.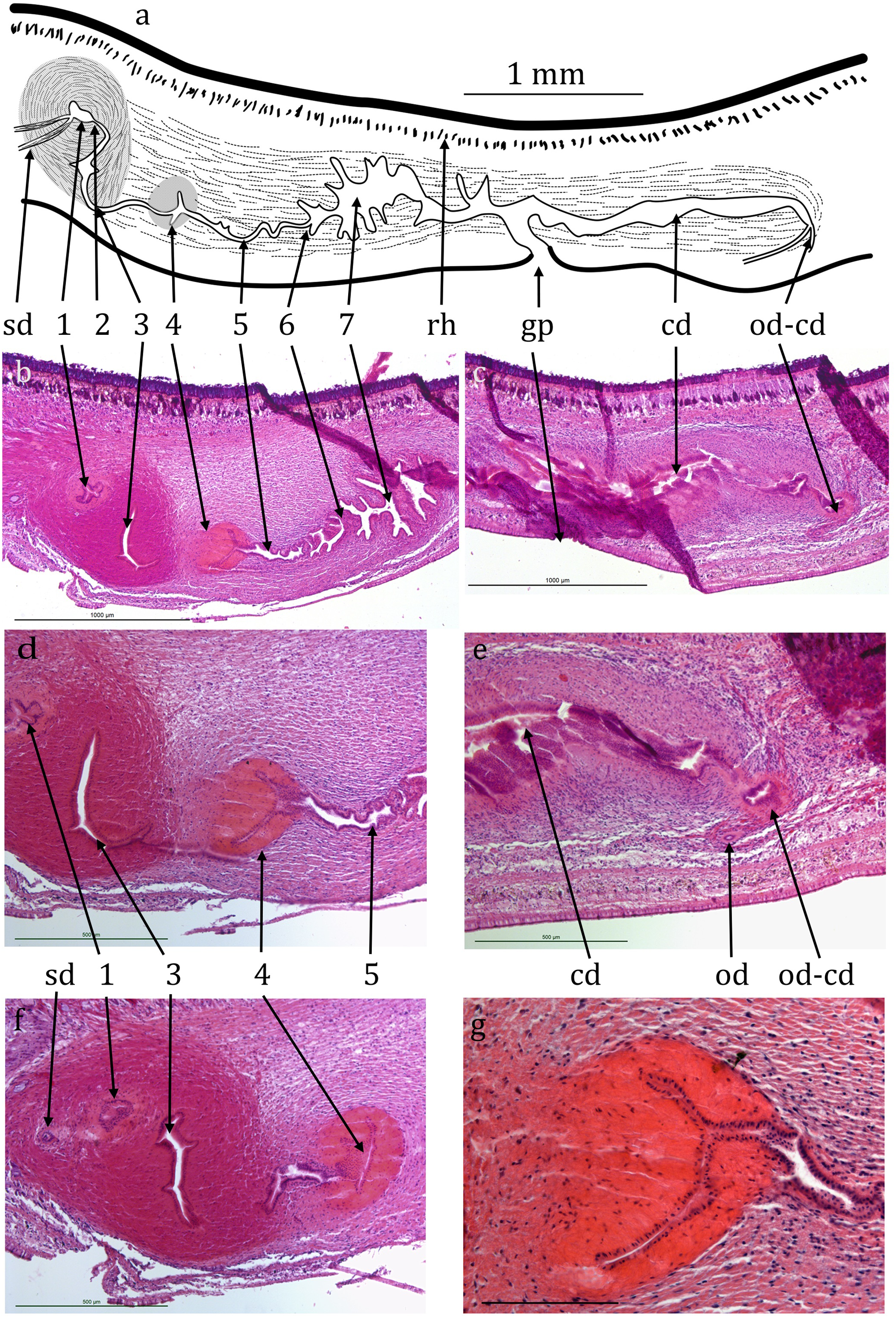{kind=link}
Testes are numerous, ventral, ovate, about 200 µm wide and 300 µm high (Figs. 4A, 4D and 4E) and run almost to the copulatory apparatus. The sperm ducts cannot be distinguished with certainty in transverse sections. They enter the anterior end of the muscular bulb of the eversible penis, widen slightly and contain small amounts of stored sperm (Fig. 4B). They separately enter the anterior end of the ejaculatory duct which is complex, long and sinuous, about 1.5 mm from its anterior end to the gonopore (Figs. 4A, 4B, 4D and 4F). It has several regions, for ease of reference they are here arbitrarily numbered 1–7 from anterior to posterior as follows. (1) A small chamber (seminal vesicle) which extends transversely through 10 × 15 µm sections, thus about 150 µm wide, the two sperm ducts entering on either lateral extremity. (2) A narrow duct extending posteriorly and turning ventrally and opening into, (3) a sinus-like duct wide laterally, 23 × 15 µm thus 345 µm wide, but only 35 µm in the antero-posterior direction. This duct initially turns ventrally then narrows and curves posteriorly to be almost U-shaped (second arm shorter). The ejaculatory duct continues into, (4) a narrow sinus-like lumen surrounded by strongly eosinophilic cells forming a structure roughly spherical in outline about 400 µm in diameter. The cells of this region appear to be elongate with nuclei mostly adjacent to the lumen (Fig. 4G). This in turn opens into, (5) a portion about 400 µm long with sinuous margins, which in turn opens via, (6) a small papilla-like opening into, (7) a longer and wider duct about 600 µm long with sinuous walls which can be considered to be the male antrum. This in turn opens to the common antrum above the gonopore.
Discussion
The previous molecular results (Álvarez-Presas et al., 2014) analyzing only Caenoplana sequences (and an outgroup) indicated that C. decolorata specimens are closely related to Caenoplana variegata (Fletcher & Hamilton, 1888) (named as C. bicolor (von Graff, 1899) in that work, see Jones et al., 2020) although without support. In the present work, the tree shows a closer relationship between C. decolorata and C. coerulea, while C. variegata is sister to the clade formed by these two species (plus some putative unknown species), which will be an expected result having into account the more similar external coloration pattern of the first two species. However, the bb values do not support the relationships among species in the present work neither and make impossible to validate this hypothesis.
The sectioned specimen has multiple eyes, ventral testes, a layer of parenchymal longitudinal muscle, stronger ventrally, a long and fairly elaborate copulatory apparatus, the ejaculatory duct particularly so, and other anatomical characters of the genus Caenoplana Moseley, 1877 as amended by Ogren & Kawakatsu (1991) and by Winsor (1991). Thus we are confident of the generic placement.
However, comparison with other Caenoplana species is problematic. Ogren & Kawakatsu (1991) list 11 species of Caenoplana each with an anatomical description. Winsor (1991) lists 19 species, seven ‘provisionally placed,’ within Caenoplana. None of those has a similar external coloration to the present specimens, and the ejaculatory duct of the present specimens is distinctly different to that of any of those 11. They also differ from C. variegata (Fletcher & Hamilton, 1888) (synonymous with C. bicolor (von Graff, 1899), see Jones et al. (2020)).
Winsor (1997) lists a further six numbered, unnamed, Caenoplana species in addition to two named species, C. coerulea coerulea (Moseley, 1877) and C. bicolor (von Graff, 1899). Winsor (1998) states that 22 Caenoplana species were present in Australia, with no other details. Presumably this total includes the six numbered, unnamed, species above. Álvarez-Presas et al. (2014) list two further unnamed Caenoplana species, one the subject of this paper. Whether either of these is similar to any of Winsor’s (1997) unnamed species is unknown.
In comparing this species to other Caenoplana species or to species placed in the collective genus Australopacifica, particular attention should be made to those with a broadly similar pigment distribution, that is those with, dorsally, a narrow mid-dorsal pale line on an otherwise uniform dark colour (any dark colour) and ventrally with a more or less uniform, but different, colour. The only two species with such a distribution are C. coerulea Moseley, 1877 and C. purpurea (Dendy, 1895).
Caenoplana coerulea Moseley, 1877, originally found in New South Wales, Australia, was described as follows: ‘entire body of a dark Prussian blue colour, somewhat lighter on the under surface … with a narrow, mesial, dorsal, longitudinal stripe of white;’ 5 cm long. Hyman (1943, 1954) and Ogren (1989) have described the anatomy of similar specimens found in the USA. This species has distinctly different coloration from the present specimens and the ejaculatory duct has a different structure (Ogren, 1989). It has subsequently been found in New Zealand (Dendy, 1895), several European countries (Álvarez-Presas et al., 2014) and North and South America (Ogren, 1989; Luis-Negrete, Brusa & Winsor, 2011).
Geoplana purpurea Dendy, 1895, originally from South Island, New Zealand, was described as follows: ‘dorsal surface rather dark reddish-purple … a very narrow median band of nearly white,’ ‘ventral surface paler purple, under a lens appearing very finely mottled in two shades;’ 3.5 cm long. Dendy (1895) comments: ‘it is perhaps doubtful whether this species ought to be separated from the Australian G. coerulea, from which it differs only in colour.’ But in the same paper Dendy also records C. coerulea. Geoplana purpurea was placed by Ogren & Kawakatsu (1991) in the collective genus Australopacifica, with the note that ‘this probably belongs to Caenoplana on basis of external similarities to Caenoplana coerulea.’ Winsor (1991) ‘provisionally placed’ it within Caenoplana. There has been no anatomical description of specimens under that species name. However, the coloration is different to the specimens from Spain and it seems unlikely that the latter are of the same species.
None of the other species listed by Ogren & Kawakatsu (1991) under Australopacifica has a colouration similar to the present species.
Thus the specimens do not match the description of any species previously described and are described as a new species, Caenoplana decolorata.
One possible confusing factor is that the colour of some species has been shown to vary over time and between individuals due to feeding (Jones et al., 2020; McDonald & Jones, 2007). Only prolonged observations on live animals before and after feeding could clarify if that might be the case with this species. Such observations would also indicate its preferred prey.
The ejaculatory duct of the new species is distinctive. The structure here numbered 4 is unlike anything present in any other described species of Caenoplana or for that matter any other terrestrial planarian. The function of this structure is not obvious; it does not appear to be either glandular or muscular.
Since at least one of the specimens was mature, it is presumed that this species reproduces by sexual reproduction, though it is entirely possible that it may also reproduce by partial fission, as in C. variegata (see Jones et al., 2020) and several other land planarian species.
This species almost certainly originates from Australia since most Caenoplana species are from there. It is presumed that it has been inadvertently transported to Spain in the course of the trade in horticultural products.