
Two new species of Tardigrada from moss cushions (Grimmia sp.) in a xerothermic habitat in northeast Tennessee (USA, North America), with the first identification of males in the genus Viridiscus
- Published
- Accepted
- Received
- Academic Editor
- Michael Wink
- Subject Areas
- Biodiversity, Ecology, Molecular Biology, Taxonomy, Zoology
- Keywords
- Dioecious, Eutardigrada, Heterotardigrada, Systematics, Taxonomy, Water bears, Morphology, Molecular technique, Parthenogenesis, Anhydrobiosis
- Copyright
- © 2020 Nelson et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2020. Two new species of Tardigrada from moss cushions (Grimmia sp.) in a xerothermic habitat in northeast Tennessee (USA, North America), with the first identification of males in the genus Viridiscus. PeerJ 8:e10251 https://doi.org/10.7717/peerj.10251
Abstract
Background
The phylum Tardigrada consists of over 1,300 species that inhabit terrestrial, freshwater and marine environments throughout the world. In terrestrial habitats they live primarily in mosses, lichens, leaf litter and soil, whereas tardigrades in freshwater and marine environments are mainly found in sediments and on aquatic plants. More than 65 species have been previously reported in the state of Tennessee, USA.
Methods
Tardigrades present in moss cushions (Grimmia sp.) collected from a xerothermic habitat on the East Tennessee State University campus, Johnson City, TN, USA, were extracted, mounted on slides, identified, and counted. Additional samples of fresh dried moss were used for integrative analyses, including morphological analysis with phase contrast (PCM) and scanning electron microscopy (SEM), as well as molecular analyses of COI, 18S rRNA, 28S rRNA, and ITS-2 of the Macrobiotus and Milnesium species.
Results
Five species were found, including two species new to science: Viridiscus miraviridis sp. nov. and Macrobiotus basiatus sp. nov. Viridiscus miraviridis sp. nov. differs from other members of the genus mainly by having a different type of dorsal cuticle and some other, more subtle, morphometric characters. In addition to the two new species, Viridiscus perviridis and Viridiscus viridissimus were present, and males of Vir. viridissimus were found for the first time, the first record of males in the genus Viridiscus. Macrobiotus basiatus sp. nov. is most similar to Macrobiotus nelsonae, but it differs from Mac. nelsonae mainly by the stylet supports being situated in a more anterior position, shorter and narrower egg processes, and a smaller number of areoles around the egg processes. Moreover, the identification of Milnesium inceptum was confirmed as the first record for the USA by analysis of COI.
Introduction
The phylum Tardigrada consists of over 1,300 species (Guidetti & Bertolani, 2005; Degma & Guidetti, 2007; Degma, Bertolani & Guidetti, 2009–2020) that inhabit terrestrial, freshwater and marine environments throughout the world. In terrestrial habitats they live primarily in mosses, lichens, leaf litter and soil, whereas in freshwater and marine environments tardigrades are found mainly in sediments and on aquatic plants (Nelson, Guidetti & Rebecchi, 2015).
Studies on the distribution of terrestrial and aquatic tardigrades have been conducted primarily in Europe and North America (see e.g., McInnes, 1994; Meyer, 2013; Kaczmarek, Michalczyk & McInnes, 2014; Kaczmarek, Michalczyk & McInnes, 2015; Kaczmarek, Michalczyk & McInnes, 2016; McInnes, Michalczyk & Kaczmarek, 2017). In addition to a few specimens reported by Riggin (1962) and Maucci (1987), surveys of the distribution of tardigrades in Tennessee have been conducted by Nelson (1975) and her students and colleagues (Kathman & Nelson, 1987; Nelson, Kincer & Williams, 1987; Nelson & McGlothlin, 1993; Nelson & McGlothlin, 1996; Kendall-Fite & Nelson, 1996; Guidetti, 1998; Bertolani, Marley & Nelson, 1999; Guidetti, Bertolani & Nelson, 1999; Nelson, Marley & Bertolani, 1999; Nelson & Adkins, 2001; Bartels & Nelson, 2006; Bartels & Nelson, 2007; Bartels & Nelson, 2012; Bartels et al., 2007; Bartels et al., 2008; Bartels et al., 2009; Bartels et al., 2011; Bartels, Nelson & Exline, 2011; Bartels et al., 2014; Bartels, Mormino & Nelson, 2017; Nelson & Bartels, 2007; Nelson & Bartels, 2013; Nelson, Bartels & Guil, 2019). Thus far for the state of Tennessee, more than 65 species have been reported (Kaczmarek, Michalczyk & McInnes, 2016; Meyer, 2013).
The genus Viridiscus was recently established by Gąsiorek & Michalczyk in Gąsiorek et al. (2019) in an integrative analysis of the “arctomys group” of Echiniscus, in which five new genera were erected. The widely distributed “Echiniscus viridis group” (now Viridiscus), with the very characteristic fully or partially green cuticle and lacking all lateral and dorsal appendages except cirrus A currently includes six species: Viridiscus clavispinosus (Fontoura, Pilato & Lisi, 2011), Viridiscus perviridis (Ramazzotti, 1959), Viridiscus rufoviridis (Du Bois Reymond Marcus, 1944), Viridiscus viridianus (Pilato, Fontoura & Lisi, 2007), Viridiscus viridis (Murray, 1910), and Viridiscus viridissimus (Péterfi, 1956). In this paper, we provide a description of a new species of Viridiscus, and the first confirmation of the presence of males in the genus.
We also provide the first record of Milnesium inceptum Morek et al., 2019 from the USA (Tennessee, Washington County), confirmed by molecular analysis. Milnesium Doyère, 1840 was originally believed to be a monospecific genus, but multiple species have been described in recent years with over 40 known species to date (Morek et al., 2020).
In addition, based on an integrative approach, we also provide a description of a new species in the genus Macrobiotus. The cosmopolitan genus Macrobiotus C.A.S. Schultze, 1834, the first described genus of tardigrades, comprises a large complex of species, some of which have been allocated to several new genera in recent years.
Materials & Methods
Sample processing
In a previous study (Nelson & Adkins, 2001), 60 samples of the moss Grimmia sp. were collected from vertical surfaces of concrete caps on six brick fence posts (10 samples per post), fully exposed to sun, wind and precipitation, located on the north side of the East Tennessee State University campus (Johnson City, Washington County, Tennessee, USA) (Figs. 1A–1C), from 14 May 1995 to 30 September 1995 (for more details see Nelson & Adkins, 2001).

Figure 1: Sampling site and type locality of Viridiscus miraviridis sp. nov. and Macrobiotus basiatus sp. nov. in moss cushions (Grimmia sp.) on the campus of East Tennessee State University, Johnson City, Tennessee.
(A) Overall view of one of the brick fence posts on the campus. (B) Close up of the concrete cap on the fence post. (C) Close up of the moss showing the cushion growth form and leaf structures. Photo credit: Diane R. Nelson.{kind=link}
The samples were soaked in tap water in a small jar for 30 min, then agitated vigorously. Afterwards, the sample was poured into a beaker through a wire screen to remove the moss. When the beaker contents settled, the top layer of water was decanted, and boiling alcohol was added to preserve the sample. The beaker contents were then poured through a 45 µm sieve. The remaining sample on the sieve was washed into a petri dish and examined for tardigrades under a stereomicroscope. All tardigrades and eggs found in each sample were extracted with an Irwin loop. To describe the two new species with an integrative approach, additional samples of freshly dried moss from the same posts were collected in 2019 and mailed to Adam Mickiewicz University, Poznań, Poland, for processing, imaging and morphometrics.
Microscopy and imaging
Specimens from the previous study were mounted on microscope slides in Hoyer’s mounting medium, with the coverslip sealed with epoxy paint when dry. All specimens and eggs were identified to species level with phase contrast light microscopy (PCM). Microphotographs of specimens from the fresh samples were obtained with Olympus BX41 associated with an ARTCAM–300Mi digital camera (Olympus Corporation, Shinjuku–ku, Japan). A total of 10 adults of Viridiscus miraviridis sp. nov. and 25 specimens and 15 eggs of Macrobiotus basiatus sp. nov. from the fresh samples were prepared for SEM imaging according to Roszkowska et al. (2018). These animals and eggs were examined under high vacuum in a Hitachi S3000N Scanning Electron Microscope.
All figures were assembled in Corel Photo-Paint 17. For deep structures that could not be fully focused in a single photograph, a series of 2–10 images were taken every ca. 0.5 µm and then assembled into a single stacked image manually in Corel Photo-Paint 17.
Morphometrics and morphological nomenclature
Structures were measured only if their orientation was suitable. Body length was measured from the anterior extremity to the end of the body, excluding the hind legs. The types of buccal-pharyngeal apparatuses and claws were classified according to Pilato & Binda (2010). The terminology used to describe the genus Viridiscus followed Gąsiorek et al. (2019). Lengths of the claw branches for Viridiscus were measured from the base of the claw to the top of the branch. The sp index, the ratio of the length of a given structure to the length of the scapular plate (scp), was calculated according to Dastych (1999). All measurements of adults and eggs of Macrobiotus were prepared according to Kaczmarek & Michalczyk (2017). Terminology describing the oral cavity armature followed Michalczyk & Kaczmarek (2003). The macroplacoid length sequence was determined according to Kaczmarek et al. (2014). The pt ratio, the length of a given structure to the length of the buccal tube expressed as a percentage (Pilato, 1981), was also calculated. The age structure of the Viridiscus populations was determined according to Bertolani et al. (1984): 1st instar, anus and gonopore absent, 2 claws per leg present; 2nd instar, anus present but gonopore absent or rudimentary, 4 claws per leg present; adult, anus and gonopore present, 4 claws per leg present.
Morphometric data were handled using the “Parachela” ver. 1.1 and “Echiniscoidea” ver. 1.1 templates available from the Tardigrada Register (Michalczyk & Kaczmarek, 2013). Tardigrade taxonomy followed Bertolani et al. (2014). Genera were abbreviated according to Perry, Miller & Kaczmarek (2019).
Genotyping
All specimens of Macrobiotus and Milnesium were preliminarily identified using light microscopy before DNA extraction. DNA was extracted from individual animals following a Chelex®100 resin (Bio-Rad) extraction method (Casquet, Thebaud & RG, 2012) modified to obtain tardigrade exoskeletons according to Kaczmarek et al. (2019). For Macrobiotus specimens, we sequenced four DNA fragments differing in mutation rates: the cytochrome oxidase subunit I (COI, mtDNA), the internal transcribed spacer (ITS-2, nDNA), the large ribosome subunit (28S rRNA, nDNA) and the small ribosome subunit (18S rRNA, nDNA). For Milnesium specimens we sequenced only the COI fragment. All fragments were amplified and sequenced with the protocols described in Kaczmarek et al. (2019). Amplification of 28S rRNA and 18S rRNA fragments was conducted in the reaction mixture with addition of 3% DMSO. Primers used for PCR, and later sequencing, with their original references, are listed in Table 1. PCR programs used for the amplification of COI, ITS-2, 28S rRNA and 18S rRNA are listed in Table 2. For analysis of COI, PCR products were purified by thermosensitive Exonuclease I and FastAP Alkaline Phosphatase (Fermentas, Thermo Scientific), whereas ITS-2, 28S rRNA and 18S rRNA (after excision from the gel) were treated with NucleoSpin Gel and PCR Clean-up Kit (MACHERY-NAGEL). For sequencing COI, ITS-2 and 28S rRNA, the same primers were used as in the amplification reaction, while a few additional primers were required to sequence the 18S rRNA (see Table 1).
| DNA fragment | Application | Direction | Code | Sequence (5′–3′) | Reference |
|---|---|---|---|---|---|
| COI | PCR & sequencing | Forward | LCO 1490 | GGTCAACAAATCATAAAGATATTGG | Folmer et al. (1994) |
| Reverse | HCO 2198 | TAAACTTCAGGGTGACCAAAAAATCA | |||
| ITS-2 | PCR & sequencing | Forward | ITS2_Eutar_Ff | CGTAACGTGAATTGCAGGAC | Stec, Kristensen & Michalczyk (2018) |
| Reverse | ITS2_Eutar_Rr | TGATATGCTTAAGTTCAGCGG | |||
| 28S rRNA | PCR & sequencing | Forward | 28SF0001 | ACCCVCYNAATTTAAGCATAT | Mironov, Dabert & Dabert (2012) |
| Reverse | 28SR0990 | CCTTGGTCCGTGTTTCAAGAC | |||
| 18S rRNA | PCR & sequencing | Forward | Euk-A | AACCTGGTTGATCCTGCCAGT | Koid et al. (2012) |
| Reverse | Euk-B | GATCCTTCTGCAGGTTCACCTAC | |||
| sequencing | Forward | fw390 | AATCAGGGTTCGATTCCGGAGA | Dabert et al. (2010) | |
| Forward | for_770 | ACTTTGAAAAAATTAGAGTGC | |||
| Reverse | rev930 | GACGGTCCAAGAATTTCAC | |||
| Reverse | rev_1460 | CATCACAGACCTGTTATTGC |
The sequences were edited and manually checked against non-conservative alignments using BioEdit, version 7.0.5. (Hall, 1999). The 18S rRNA sequence was assembled using the CAP contig assembly program implemented in BioEdit. All sequences were submitted to GenBank (see ‘Results’ section).
We were not able to obtain DNA from the Viridiscus specimens since more than one morphologically very similar species of Viridiscus were present and insufficient numbers were present in the new samples from the study sites.
Comparative molecular analysis
The obtained sequences were analysed by Standard Nucleotide BLAST to confirm their uniqueness. Then, a comparison was performed with the COI sequence of the genus Milnesium, as well as COI, ITS-2, 18S rRNA, and 28S rRNA sequences of the genus Macrobiotus, deposited in GenBank, using only the sequences of good quality and length. All sequences were aligned with the ClustalW Multiple Alignment tool (Thompson, Higgins & Gibson, 1994) implemented in BioEdit. Alignment sequences were trimmed to 759, 678, 276 and 518 bp for 18S rRNA (21 sequences, 17 species), 28S rRNA (18 sequences, 11 species), ITS2 (21 sequences, 12 species) and COI (32 haplotypes, 59 sequences, 16 species) barcodes, respectively, and calculation for the uncorrected p-distances was performed using MEGA X (Kumar et al., 2018). Uncorrected pairwise distances are provided in Table S1. Table S2 provides the name, accession number and link for the third-party database where the sequences have been deposited.
| COI and ITS-2 | 28S rRNA | 18S rRNA | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Step | Cycles | Time [min.:sec.] | Temp. [°C] | Cycles | Time [min:sec] | Temp. [°C] | Cycles | Time [min:sec] | Temp. [°C] |
| initial denaturation | – | 03:00 | 98 | – | 05:00 | 98 | – | 05:00 | 98 |
| denaturation | 40 | 00:15 | 98 | 40 | 00:30 | 98 | 40 | 00:30 | 98 |
| annealing | 00:30 | 50 | 01:00 | 50 | 01:00 | 60 | |||
| extension | 00:30 | 72 | 01:00 | 72 | 01:30 | 72 | |||
| final extension | – | 07:00 | 72 | – | 07:00 | 72 | – | 07:00 | 72 |
Comparative material
For comparison with the new species, the following specimens were used: Vir. viridis collected in Hawaii (Schuster collection at the Bohart Museum, University of California-Davis, USA); Vir. viridianus (paratype 060511PF) collected in Azores Is. (Fontoura Collection, Department of Zoology and Anthropology, University of Porto, Portugal); Vir. rufoviridis collected in Ecuador (slides 13775, 13779, 13814, 13817); Vir. viridissimus collected in Johnson City, TN, USA (slides 7731, 7735, 7843); Vir. viridis collected in Ecuador (slide 13821); Vir. perviridis collected in Johnson City, TN, USA (slide 7744) and in Italy (slide 4564) (Maucci collection at the Natural History Museum of Verona, Italy); and holotype and paratypes of Macrobiotus nelsonae (Guidetti, 1998) collected on Roan Mountain, Tennessee (slides 5N05A-S10) (Bertolani Collection, Department of Life Science, University of Modena and Reggio Emilia, Italy).
In addition to the key to Viridiscus in Fontoura, Pilato & Lisi (2011), the following original descriptions or redescriptions, as well as more recent papers, were used: Murray (1910), Du Bois Reymond Marcus (1944), Péterfi (1956), Ramazzotti (1959), Pilato, Fontoura & Lisi (2007); Pilato, Fontoura & Lisi (2008), and Morek et al. (2019).
For the differential diagnosis of the new Macrobiotus species, the following original descriptions or redescriptions were used: Maucci (1954), Guidetti (1998), Binda et al. (2001) and Meyer et al. (2017).
For the correct identification of Mil. inceptum, we used the original description in Morek et al. (2019).
The electronic version of this article in Portable Document Format (PDF) will represent a published work according to the International Commission on Zoological Nomenclature (ICZN), and hence the new names contained in the electronic version are effectively published under that Code from the electronic edition alone. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix http://zoobank.org/ . The LSID for this publication is: urn:lsid:zoobank.org:pub:7E91563A-395E-4B52-8AD1-009CFABBD472. The online version of this work is archived and available from the following digital repositories: PeerJ, PubMed Central and CLOCKSS.
Results
Five tardigrade species from three genera were found in the present study: Mil. inceptum (Eutardigrada, Apochela), Macrobiotus basiatus sp. nov. (Eutardigrada, Parachela), and three species of Heterotardigrada, Vir. miraviridis sp. nov., Vir. perviridis, and Vir. viridissimus.
In terms of relative abundance, Vir. viridissimus was the most numerous species, followed by Mil. inceptum; both species were present on all six posts. The other three species were much less numerous with Macrobiotus basiatus sp. nov. found on four of the posts, whereas Vir. perviridis and Vir. miraviridis sp. nov. were present on only two of the posts.
Taxonomic account
| Phylum: Tardigrada Doyère, 1840 |
| Class: Heterotardigrada Marcus, 1927 |
| Family: Echiniscidae Thulin, 1928 |
| Genus:Viridiscus Gąsiorek & Michalczyk, 2019, in Gąsiorek et al., 2019 |
1. Viridiscus miraviridis sp. nov. Genus name: urn:lsid:zoobank.org:act:2FCD58BD-5429-451A-BE2D-D6BA33C1ED31, Species name: urn:lsid:zoobank.org:act:0A9781C0-8F03-4A68-BE35-46DDB066E246. Publication LSID: urn:lsid:zoobank.org:pub:7E91563A-395E-4B52-8AD1-009CFABBD472.
| CHARACTER | N | RANGE | MEAN | SD | Holotype | ||||
|---|---|---|---|---|---|---|---|---|---|
| μ ma | sp | μ ma | sp | μ ma | sp | μ ma | sp | ||
| Body length | 30 | 133–247 | – | 200 | – | 29 | 14 | 191 | – |
| Scapular plate length | 30 | 30.1–53.3 | – | 43.8 | – | 6.2 | – | 42.7 | – |
| Head appendages lengths | |||||||||
| Cirrus internus | 30 | 8.9–17.8 | 25.4–34.4 | 13.5 | 30.7 | 2.3 | 2.3 | 13.5 | 31.6 |
| Cephalic papilla | 29 | 4.2–7.6 | 12.4–15.9 | 6.1 | 14.0 | 0.9 | 0.8 | 6.1 | 14.3 |
| Cirrus externus | 29 | 10.0–21.0 | 30.5–40.6 | 15.0 | 34.4 | 2.5 | 2.3 | 14.9 | 34.9 |
| Clava | 30 | 3.8–7.2 | 12.6–16.4 | 5.9 | 13.6 | 0.7 | 0.9 | 5.8 | 13.6 |
| Cirrus A | 27 | 31.2–65.4 | 103.7–126.5 | 50.9 | 114.9 | 7.8 | 6.2 | 50.3 | 117.8 |
| Cirrus A/Body length ratio | 27 | 21%–30% | – | 25% | – | 2% | – | 26% | – |
| Cirrus int/ext length ratio | 29 | 77%–96% | – | 89% | – | 4% | – | 91% | – |
| Spine on leg I length | 29 | 1.8–3.5 | 5.5–7.1 | 2.8 | 6.3 | 0.5 | 0.4 | 2.8 | 6.6 |
| Papilla on leg IV length | 23 | 3.0–4.7 | 8.3–10.4 | 4.0 | 9.1 | 0.4 | 0.5 | 3.9 | 9.1 |
| Number of teeth on the collar | 25 | 8–11 | – | 9.4 | – | 0.7 | – | 10 | – |
| Notch length | 24 | 11.4–19.3 | 33.9–39.9 | 16.1 | 36.7 | 2.1 | 1.8 | 16.6 | 38.9 |
| Claw 1 lengths | |||||||||
| Branch | 24 | 10.0–17.9 | 29.9–35.6 | 14.1 | 32.3 | 1.9 | 1.6 | 14.6 | 34.2 |
| Spur length | 10 | 1.1–1.9 | 2.7–3.6 | 1.4 | 3.2 | 0.3 | 0.3 | 1.3 | 3.0 |
| Spur/branch length ratio | 10 | 9%–11% | – | 10% | – | 1% | – | 0 | – |
| Claw 2 lengths | |||||||||
| Branch | 24 | 9.0–15.4 | 27.8–34.3 | 13.1 | 30.7 | 1.7 | 1.8 | 13.2 | 30.9 |
| Spur length | 12 | 0.9–1.7 | 2.9 – 4.2 | 1.5 | 3.3 | 0.2 | 0.4 | ? | ? |
| Spur/branch length ratio | 12 | 9%–13% | – | 11% | – | 1% | – | ? | – |
| Claw 3 lengths | |||||||||
| Branch | 20 | 9.8–16.6 | 27.9–33.5 | 13.4 | 30.5 | 1.6 | 1.6 | 13.6 | 31.9 |
| Spur length | 9 | 1.2–1.5 | 2.5 – 3.8 | 1.4 | 3.0 | 0.1 | 0.3 | 1.3 | 3.0 |
| Spur/branch length ratio | 9 | 9%–12% | – | 10% | – | 1% | – | 0 | – |
| Claw 4 lengths | |||||||||
| Branch | 25 | 11.4–19.7 | 34.3–40.7 | 16.2 | 37.1 | 2.1 | 1.7 | 15.1 | 35.4 |
| Spur length | 20 | 1.2–2.1 | 3.2 – 4.7 | 1.7 | 3.9 | 0.2 | 0.4 | 1.7 | 4.0 |
| Spur/branch length ratio | 20 | 9%–13% | – | 11% | – | 1% | – | 0 | – |
Notes:
- N
-
number of specimens/structures measured, RANGE refers to the smallest and the largest structure among all measured specimens
- SD
-
standard deviation
- ?
-
trait oriented unsuitably for measurement
- sp
-
ratio of the length of a given structure to the length of the scapular plate
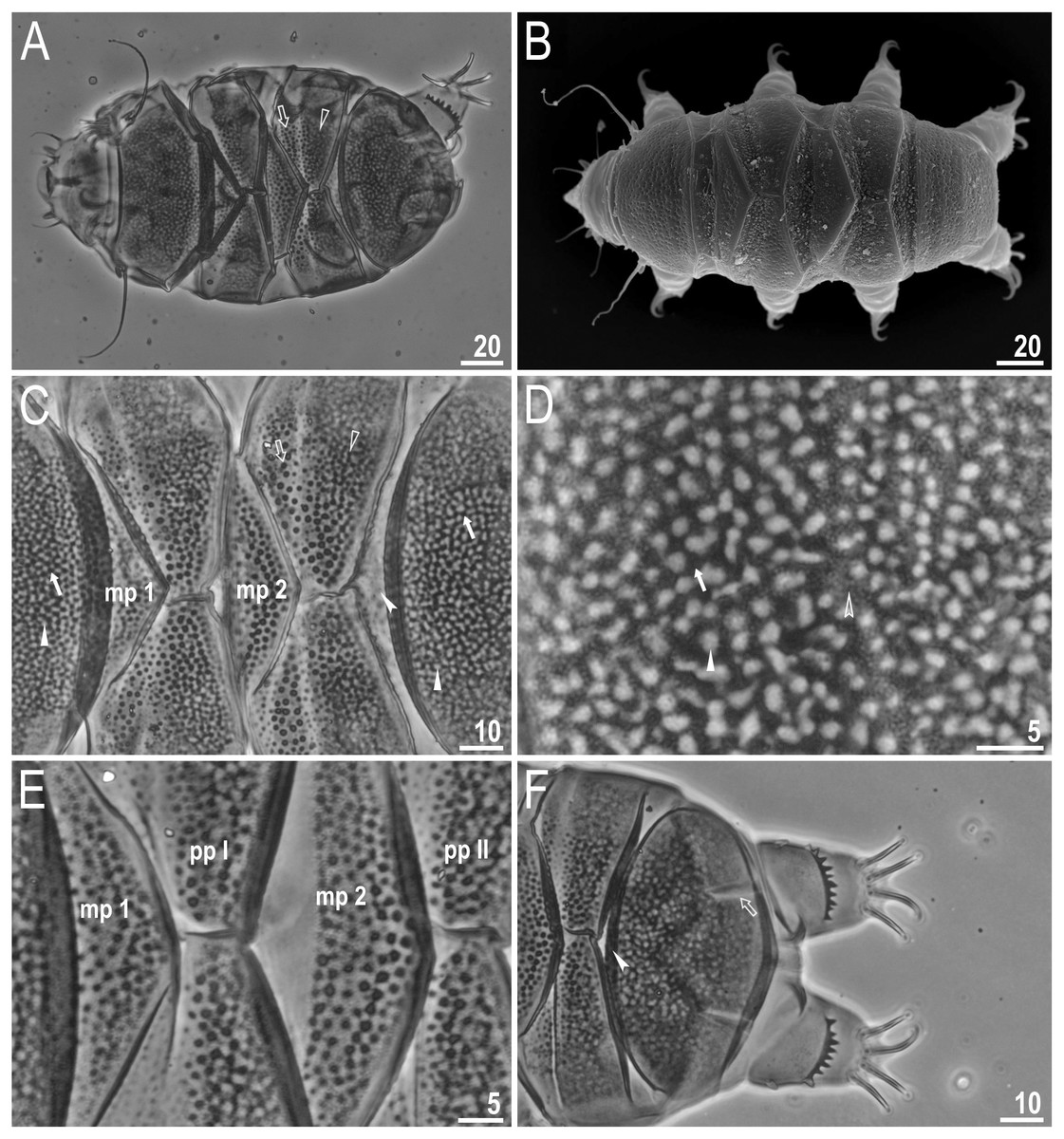
Figure 2: Viridiscus miraviridis sp. nov.
(A) Habitus, dorsoventral assembled image, empty arrow indicates the anterior part of the paired plates, empty arrowhead indicates the posterior part of the paired plates (holotype, PCM). (B) Dorsal view (paratype, SEM). (C) Close up of the dorsal cuticle, empty arrow indicates the anterior part of the paired plates with large, dark unconnected granules, empty arrowhead indicates the posterior part of the paired plates with a mesh of irregular dark granules connected by dark lines, filled arrow indicates large, irregular dark granules, filled arrowhead indicates irregular light depressions between a mesh of dark granules, filled indented arrowhead indicates the absence of median plate 3, mp 1 and mp 2 indicate median plate 1 and 2, respectively (paratype, PCM). (D) Close up of the terminal plate, filled arrow indicates large, irregular dark granules, empty indented arrowhead indicates a dense background of fine dots, filled arrowhead indicates irregular light depressions between a mesh of dark granules (paratype, PCM). (E) Close up of the dorsal cuticle, mp 1 and mp 2 indicate median plate 1 and 2, respectively, pp I and pp II indicate paired plates I and II, respectively (paratype, PCM). (F) Terminal plate with two notches (empty arrow), filled indented arrowhead indicates the absence of median plate 3 (paratype, PCM). Scale bars in micrometres [µm]. Photo credit: Milena Roszkowska.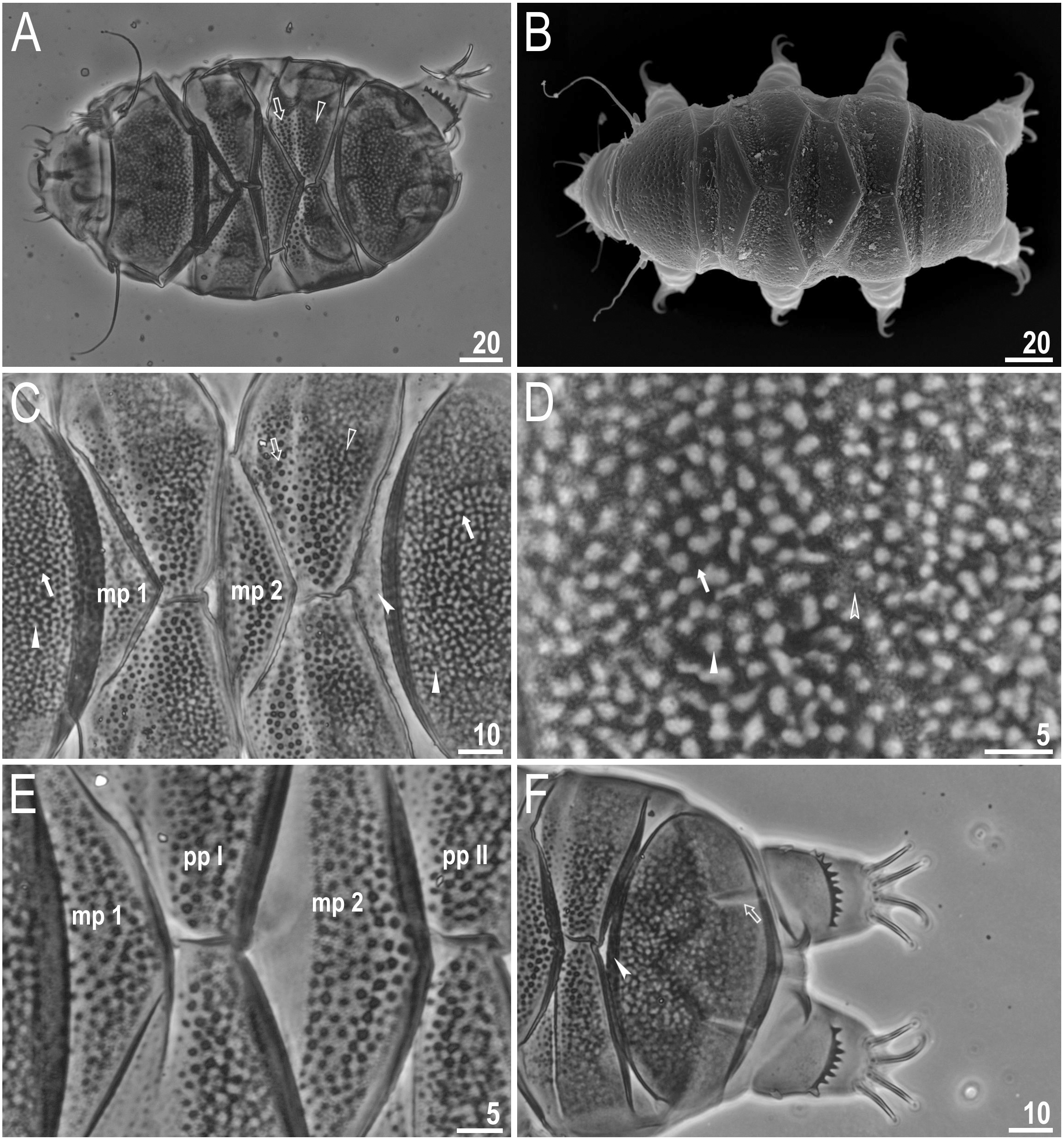{kind=link}
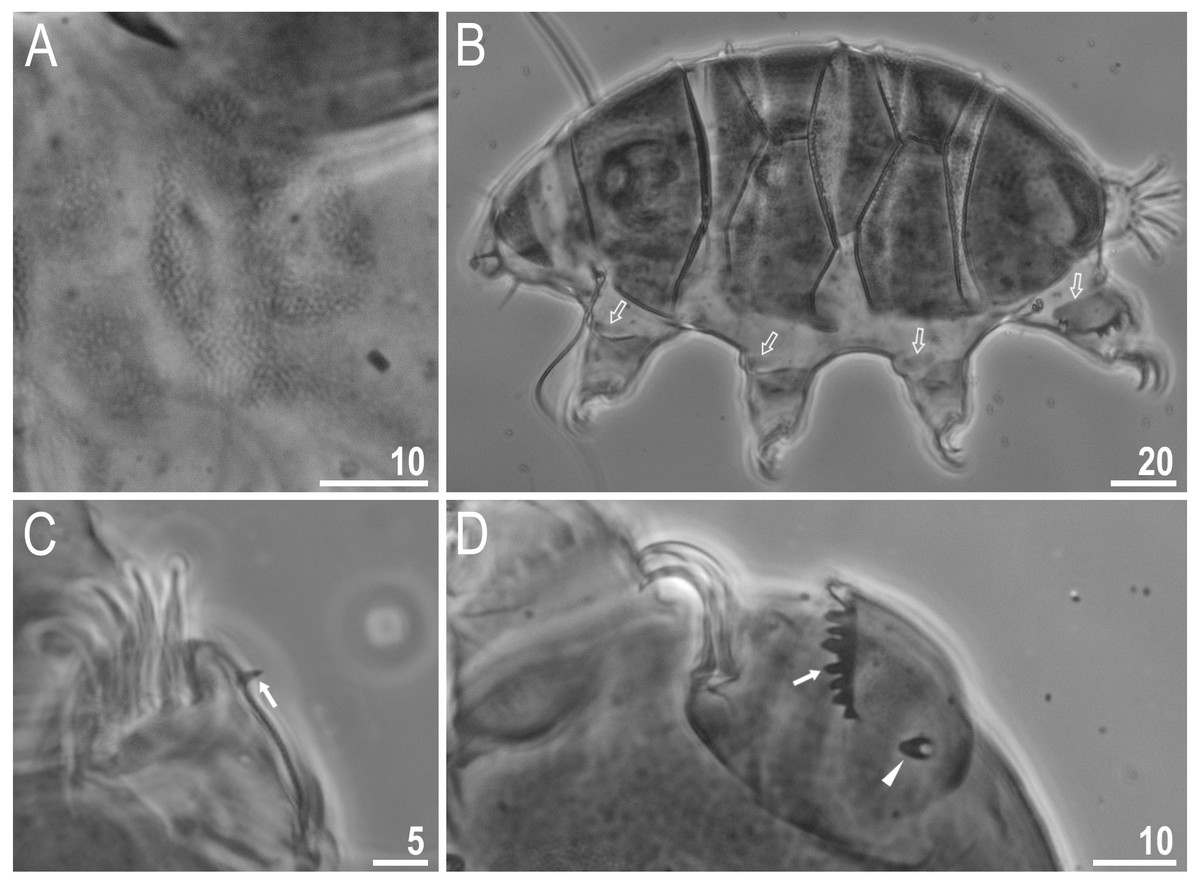
Figure 3: Viridiscus miraviridis sp. nov.
(A) Ventral cuticle with tiny, regular granulation (paratype, PCM). (B) Animal in dorsolateral view, empty arrows indicate the narrow bands of granules on all legs (paratype, PCM). (C) Leg I, filled arrow indicates the spine on leg I (holotype, PCM). (D) Leg IV, filled arrowhead indicates the papilla on leg IV, filled arrow indicates the dentate collar (holotype, PCM). Scale bars in micrometres [µm]. Photo credit: Milena Roszkowska.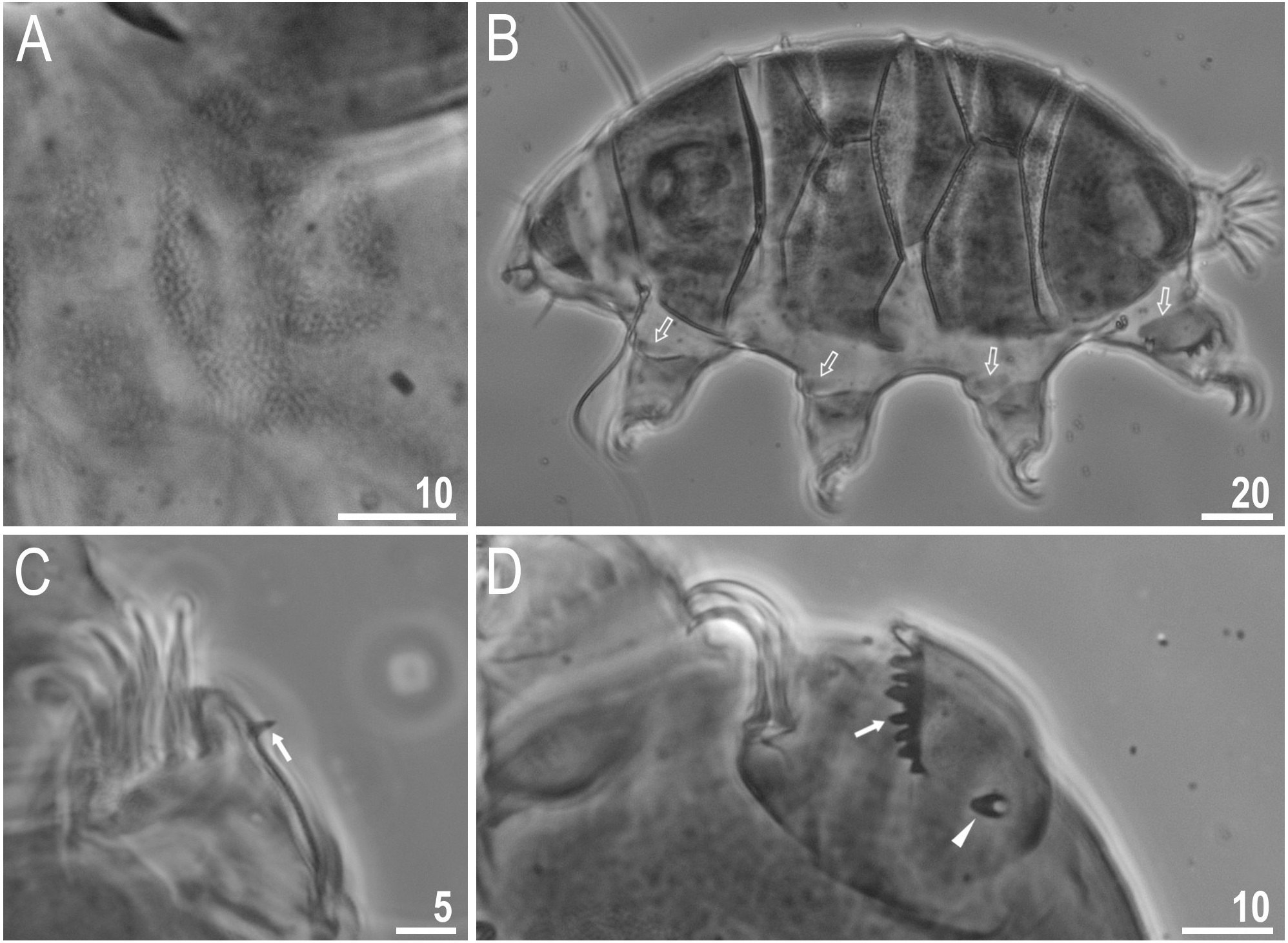{kind=link}
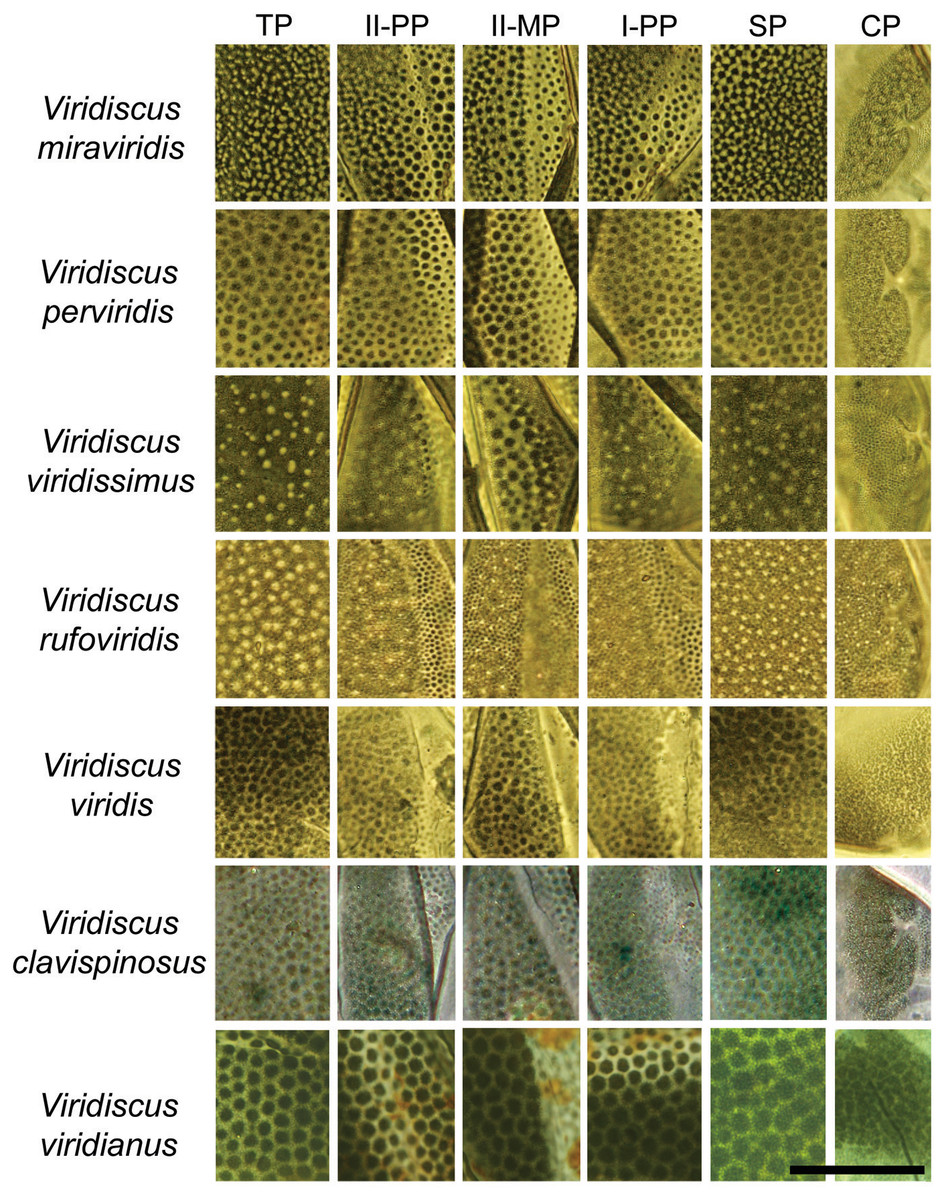
Figure 4: Comparison of the types of dorsal sculpture in the seven Viridiscus species (PCM).
Vir. miraviridis sp. nov., Vir. perviridis, Vir. viridissimus, Vir. rufoviridis, Vir. viridis, Vir. clavispinosus, and Vir. viridianus are illustrated (green filter enhanced contrast/colour). The characters used in the comparison are: TP, terminal plate. II-PP, second paired plate. II-MP, second median plate. I-PP, first paired plate. SP, scapular plate. CP, cephalic plate. Scale bar = 20 µm. Photo credit: Roberto Guidetti, except Vir. clavispinosus by Giovanni Pilato.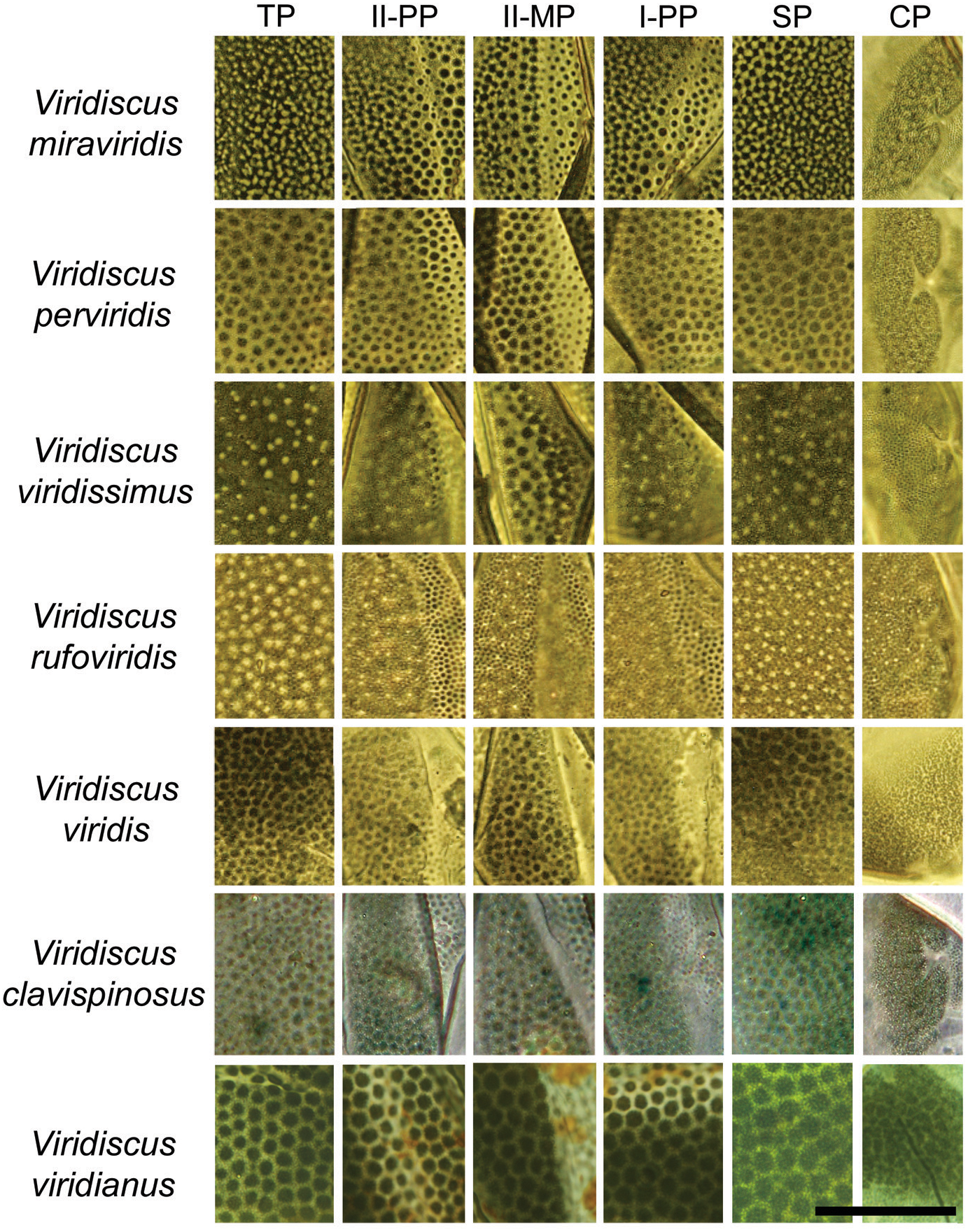{kind=link}
Type locality: 36°18′N, 82°22′W, 517 m asl; north side of the East Tennessee State University campus, Johnson City, Washington County, TN, USA, moss (Grimmia sp.) on concrete caps of brick fence posts; xerothermic habitat fully exposed to sun and wind.
Etymology: The species name is based on mira (L.) - wonderful, remarkable and viridis (L.) - green, which refers to the beautiful green color of the cuticle.
Type material: Holotype and 49 paratypes: 3 specimens of the 2nd instar; 20 specimens of adults) identified as females; and 27 individuals whose gender was not distinguished due to poor position on the slide or indistinct gonopore (i.e., as small, round circles that could not be definitely associated with a gender). No 1st instar.
Type depositories: The holotype and 9 paratypes are deposited in the National Museum of Natural History, Smithsonian Institution, Washington, DC, USA. (holotype and 4 paratypes, slide number USNM 1622558; 5 paratypes, slide number USNM 1622559). Additional paratypes are deposited in the Nelson Collection (28 slides), East Tennessee State University, Johnson City, TN, USA.; Bertolani Collection (2 slides with 7 specimens), University of Modena and Reggio Emilia, Modena, Italy; and Kaczmarek Collection (10 slides), Adam Mickiewicz University, Poznań, Poland.
Description of the new species
Animals (measurements and statistics in Table 3)
Females. Body (Figs. 2A–2B) green, eyes absent or not visible after mounted in Hoyer’s. Except for the head appendages (internal and external cirri and drop-shaped cephalic papillae), only lateral cirrus A is present (with finger-like clava near the base) (Figs. 2A–2B). Gonopore rosette-shaped, anterior to the anus.
Cuticle on dorsal plates in general appears “two-layered,” constituted by a surface with shallow depressions over a uniform background of small granules (dark dots) (Figs. 2C–2F, 3). Head plate with very light mesh sculpture over a background of fine dots. On the scapular plate, terminal plate, and posterior parts of paired plates, there are large irregular dark granules and shallow depressions (in PCM) (Figs. 2C–2D, filled arrows) over a dense background of fine dots (Fig. 2D, empty indented arrowhead). These granules appear connected by dark lines, forming a lattice-like network interspersed with irregular light depressions (Figs. 2C–2D, filled arrowheads). Meshes are more irregular on the terminal plate (Fig. 2D). Dorsal plates are well developed. Head and scapular plates are not faceted. Paired plates I and II are divided into two parts, with anterior portion having a background of fine dots and large dark granules that are not connected (Figs. 2A, 2C, empty arrow), and the posterior portion with the sculpture as described above (Figs. 2A, 2C, empty arrowhead). Additionally, on lateral sides of the anterior parts of paired plates I and II, a small area without granules is present (but with background of fine dots). Median plates 1 and 2 are divided into two parts with different types of sculpture (Figs. 2C, 2E). Median plate 1 and 2 have cuticular pattern similar to the scapular plate, terminal plate, and posterior parts of paired plates. Median plate 3 is absent; instead there are dark, round granules over a very poorly visible background of fine dots (Figs. 2C, 2F, filled indented arrowhead). Terminal plate has two notches (Fig. 2F, empty arrow). Ventral plates are absent, but the cuticle has a tiny, regular granulation caused by dense cuticular pillars (Fig. 3A).
All legs have a narrow band of very fine granules (Fig. 3B, empty arrows). Dentate collar on legs IV has fine granulation and 8–14 teeth (teeth usually have only a single point and are slightly irregular) (Figs. 2F, 3D). Triangular spine on leg I (Fig. 3C, filled arrow) and finger-like papilla on leg IV are present (Fig. 3D, filled arrowhead). External claws of all legs are smooth, internal have spurs directed downwards; spurs are larger on the claws on the hind legs.
Males and eggs unknown.
Differential diagnosis
Based on the specific morphology of the cuticle on the dorsal plates, the new species differs from all other species in the viridis group (Fig. 3). More specifically Vir. miraviridis sp. nov. differs from:
1. Viridiscus clavispinosus, known only from its type locality in Republic of Cape Verde (Santo Antão Island off West Africa in the Atlantic Ocean) (Fontoura, Pilato & Lisi, 2011), by: different type of dorsal cuticle (in general dorsal granules appear connected by dark lines in PCM, forming a lattice-like network interspersed with irregular light depressions in the new species vs slightly raised dark tubercles that are not connected in Vir. clavispinosus); different shape of clava (finger-like clava in the new species vs spine-like clava in Vir. clavispinosus); area between paired plates II and terminal plate is sculptured vs smooth in Vir. clavispinosus; slightly smaller sp index of the clava (12.6–16.4 in the new species vs 16.5–18.1 in Vir. clavispinosus); larger sp index of cirrus A (103.7–126.5 in the new species vs 54.5–66.4 in Vir. clavispinosus); and larger cirrus A/body length ratio (21–30% in the new species vs 10.9–13.4% in Vir. clavispinosus).
2. Viridiscus perviridis, known from Italy and the USA (Ramazzotti, 1959; Kaczmarek, Michalczyk & McInnes, 2016), by: different type of dorsal cuticle (in general dorsal granules appear connected by dark lines in PCM, forming a lattice-like network interspersed with irregular light depressions in the new species vs polygonal tubercles not connected in Vir. perviridis); and shorter cirrus A (31.2–65.4 µm in the new species vs 114.0–170.0 µm in Vir. perviridis).
3. Viridiscus rufoviridis, known from Argentina and Brazil (Kaczmarek, Michalczyk & McInnes, 2015), by: different type of dorsal cuticle (in general dorsal granules appear connected by dark lines in PCM, forming a lattice-like network interspersed with irregular light depressions in the new species vs large dorsal granules absent in Vir. rufoviridis); entire body green (only caudal region of the body of green colour in Vir. rufoviridis); median plate 3 absent; terminal plate not faceted; and presence of spurs on internal claws.
4. Viridiscus viridianus, known from the Azores Islands (Portugal) and the USA (Pilato, Fontoura & Lisi, 2007), by: different type of dorsal cuticle (in general dorsal granules appear connected by dark lines in PCM, forming a lattice-like network interspersed with irregular light depressions in the new species vs polygonal tubercles not connected in Vir. viridianus); and cirrus A longer than scapular plate (cirrus A shorter than scapular plate in Vir. viridianus). Viridiscus viridianus from the Azores Islands was reported to be dark grey in colour (Fontoura, 1985), rather than green as in the species from the USA and the other species of Viridiscus.
5. Viridiscus viridis, with confirmed localities only from the USA (Hawaiian Islands) (Pilato, Fontoura & Lisi, 2008), by: different type of dorsal cuticle (in general dorsal granules appear connected by dark lines in PCM, forming a lattice-like network interspersed with irregular light depressions in the new species vs polygonal tubercles not connected in Vir. viridis); longer cirrus A (31.2–65.4 µm in the new species vs 16.3–18.2 µm in Vir. viridis); larger sp index of cirrus A (103.7–126.5 in the new species vs 36.1–43.5 in Vir. viridis); and larger cirrus A/body length ratio (21–30% in the new species vs 7.4–9.6% in Vir. viridis).
6. Viridiscus viridissimus, known from Romania, the USA and Venezuela (Péterfi, 1956; Kaczmarek, Michalczyk & McInnes, 2015; Kaczmarek, Michalczyk & McInnes, 2016), by: different type of dorsal cuticle (in general dorsal granules appear connected by dark lines in PCM, forming a lattice-like network interspersed with irregular light depressions in the new species vs large dorsal granules absent in Vir. viridissimus) and shorter cirrus A (cirrus A/body length ratio is 21–30% (31.2–65.4 µm) in the new species vs 36% (85.0 µm in specimen 234.0 µm long) in Vir. viridissimus).
2. Viridiscus perviridis Ramazzotti, 1959
Type locality: Italy
Material examined: 137 specimens.
Remarks: Viridiscus perviridis has previously been reported from Italy and Tennessee (USA) (Ramazzotti, 1959; Maucci, 1987; McInnes, 1994; Nelson & Bartels, 2007; Kaczmarek, Michalczyk & McInnes, 2016).
No males were identified in this study. Based on the presence or absence of the anus and gonopore, the age structure of the population was as follows: 20, 1st instar; 22, 2nd instar; and 35, adults. For 60 individuals, 2nd instars and adults could not be distinguished.
3. Viridiscus viridissimusPéterfi, 1956
Type locality: Romania
Material examined: 2,165 specimens.
Remarks: Viridiscus viridissimus has previously been reported from Europe (Romania), North America (Alabama, North Carolina, and Tennessee; USA) and South America (Venezuela) (Péterfi, 1956; Word, Folkerts & Mason, 1976; Nelson, 1979; Grigarick, Schuster & Nelson, 1983; Maucci, 1987; Nelson, Kincer & Williams, 1987; Dewel, Dewel & Nelson, 1992; Dewel, Dewel & Roush, 1992; McInnes, 1994; Dewel & Dewel, 1996; Nelson & Adkins, 2001; Dewel & Eibye-Jacobsen, 2006; Kaczmarek, Michalczyk & McInnes, 2015; Kaczmarek, Michalczyk & McInnes, 2016).
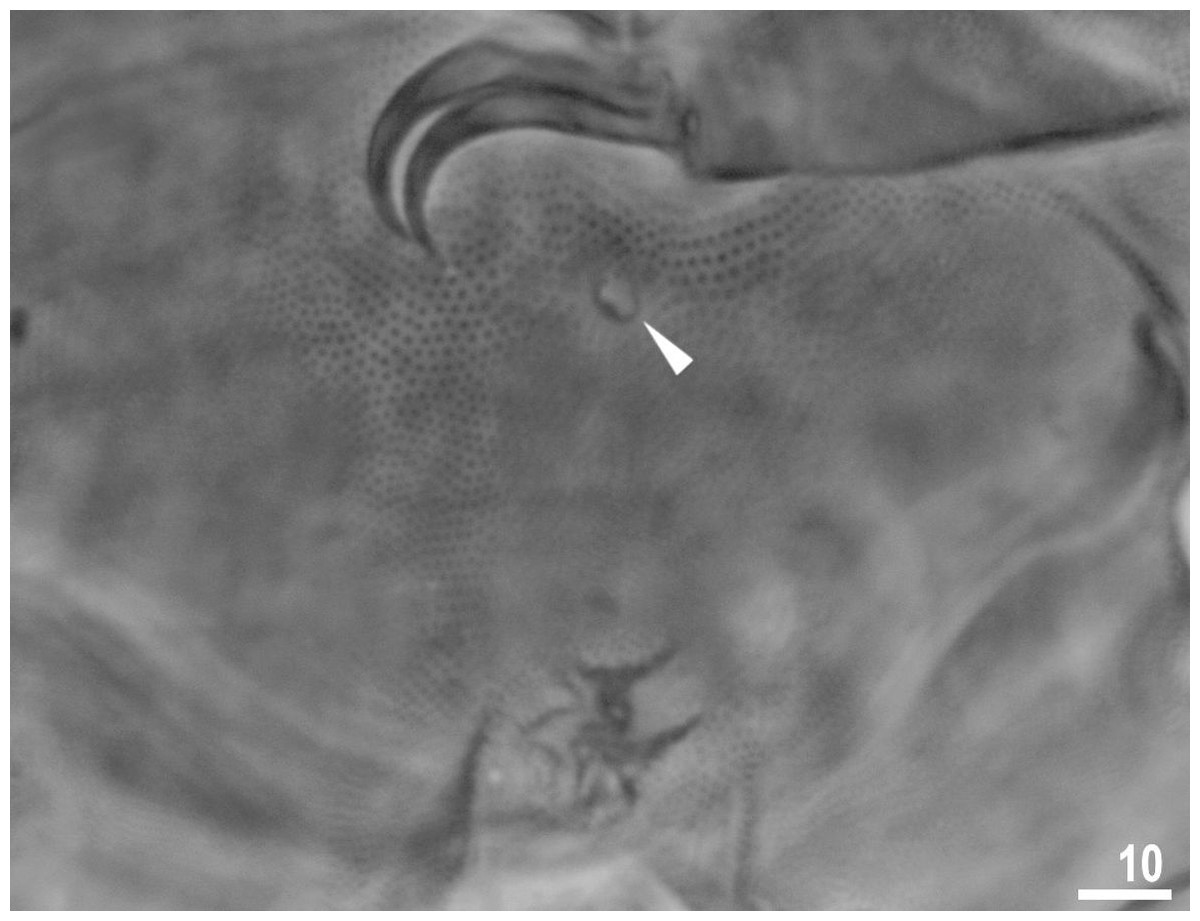
Figure 5: Male gonopore (filled arrowhead) of Viridiscus viridissimus.
Scale bars in micrometres [µm]. Photo credit: Milena Roszkowska.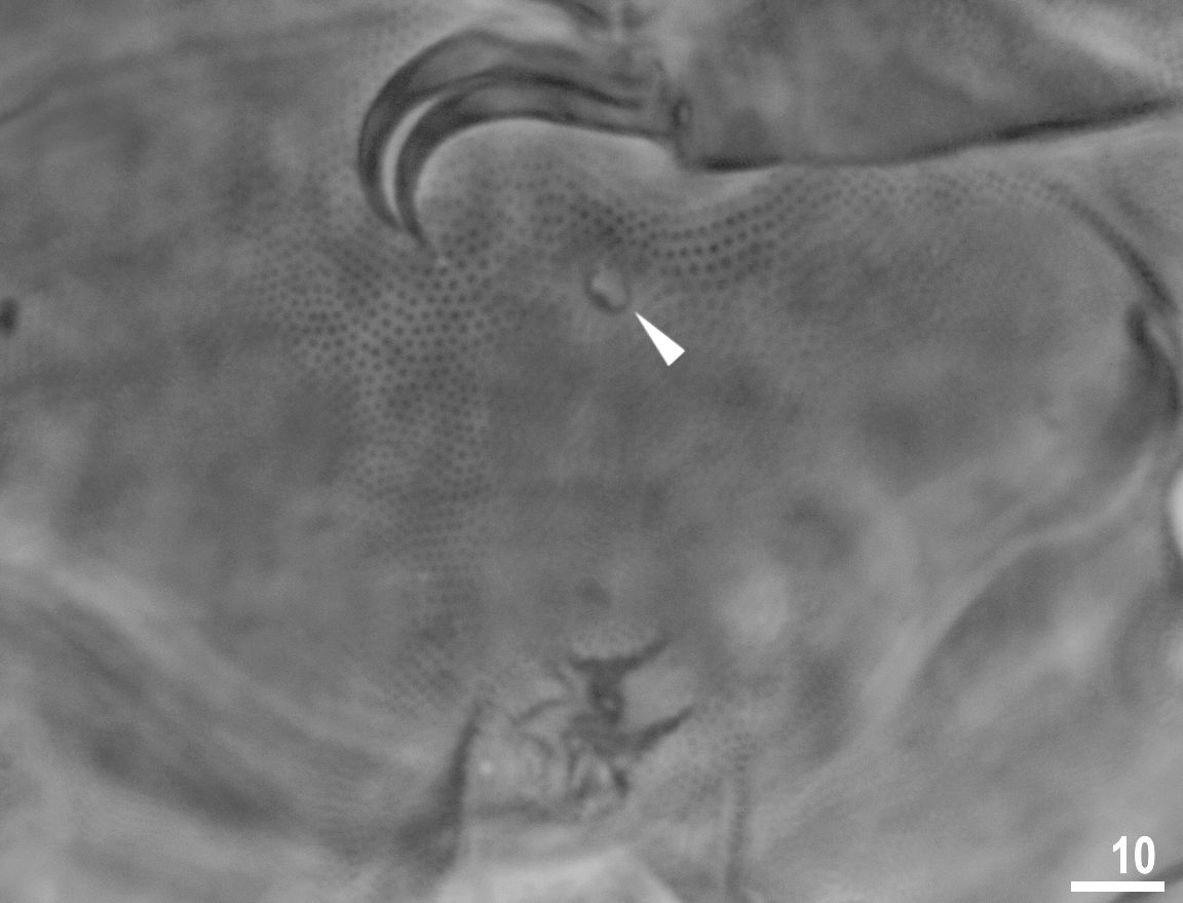{kind=link}
Males were present, but rare. The ratio of males to females for individuals whose gender could definitely be determined was 0.04 (29 males to 653 females). This is the first report of males in the genus Viridiscus. The male gonopore (Fig. 5) is oval, slightly raised, and located more posterior than the female gonopore, which has the typical rosette-shape.
The number of individuals that could be determined in each age class was as follows: 178, 1st instar; 299, 2nd instar; and 682, adults. Distinction between 2nd instar and adults could not be made for 1006 individuals due to the position of the gonopore (e.g., hidden by a leg) or due to the immaturity of the animal.
| Class: Eutardigrada Richters, 1926 |
| Order: Apochela Schuster et al., 1980 |
| Family: Milnesiidae Ramazzotti, 1962 |
| Genus:MilnesiumDoyère, 1840 |
4. Milnesium inceptumMorek et al., 2019
Type locality: Germany
Material examined: Total specimens: 1,164; sexually dimorphic males, 126.
Remarks: A pseudocryptic species, morphologically extremely similar to Milnesium alpigenum (Ehrenberg, 1853) but clearly different based on molecular data (Morek et al., 2019). This is a first record of this species from the USA. No males of Mil. inceptum were found by Morek et al. (2019) in populations from Bulgaria, Germany or Switzerland, and culturing of isolated virgin females confirmed that the type population is parthenogenetic. However, males were found to appear spontaneously in an otherwise parthenogenetic culture of the Japanese strain (Suzuki, 2008), suggesting that the species is facultatively parthenogenetic with males appearing only occasionally (Morek et al., 2019). Mature males have modified claws only on the first pair of legs, with the secondary (basal) branch of both claws shaped like a robust hook bearing a small spur (Rebecchi & Nelson, 1998).
DNA sequences
A good quality sequence for COI described here is accessible via GenBank accession number MT502117. Since the COI sequence was almost identical [only two nucleotide substitutions and p-distance 0.3% with sequence MK628723.1 (Morek et al., 2019)] to Mil. inceptum, we did not sequence 28S rRNA or ITS-2.
Class: Eutardigrada Richters, 1926
Order: Parachela Schuster et al., 1980
Superfamily: Macrobiotoidea Thulin, 1928
Family: Macrobiotidae Thulin, 1928
Genus:Macrobiotus C.A.S. Schultze, 1834
Macrobiotus basiatussp. nov. Genus name: urn:lsid:zoobank.org:act:42DFC178-D1BB-48EC-81F6-5B6BE3539AF2, Species name: urn:lsid:zoobank.org:act:95167BDA-B668-4D49-BFD2-1B314080F2F5. Publication LSID: urn:lsid:zoobank.org:pub:7E91563A-395E-4B52-8AD1-009CFABBD472.
| CHARACTER | N | RANGE | MEAN | SD | Holotype | ||||
|---|---|---|---|---|---|---|---|---|---|
| µm | pt | µm | pt | µm | pt | µm | pt | ||
| Body length | 30 | 276–540 | – | 430 | – | 80 | – | 461 | – |
| Buccalpharyngeal tube | |||||||||
| Buccal tube length | 30 | 29.3–52.6 | – | 44.4 | – | 5.8 | – | 47.6 | – |
| Stylet support insertion point | 30 | 22.7–40.7 | 74.4–78.9 | 34.5 | 77.8 | 4.5 | 1.1 | 35.7 | 75.0 |
| Buccal tube external width | 30 | 4.6–10.4 | 15.0–21.0 | 8.1 | 18.2 | 1.5 | 1.8 | 7.3 | 15.3 |
| Buccal tube internal width | 28 | 3.5–8.0 | 11.3–17.4 | 6.3 | 14.3 | 1.2 | 1.6 | 5.8 | 12.2 |
| Ventral lamina length | 29 | 19.6–31.5 | 55.7–63.7 | 26.9 | 60.0 | 3.0 | 2.0 | 27.3 | 57.4 |
| Placoid lengths | |||||||||
| Macroplacoid 1 | 30 | 7.3–17.5 | 24.6–35.0 | 13.2 | 29.6 | 2.6 | 2.8 | 13.6 | 28.6 |
| Macroplacoid 2 | 30 | 4.8–11.0 | 14.2 – 22.7 | 8.2 | 18.4 | 1.7 | 2.2 | 8.5 | 17.9 |
| Microplacoid | 30 | 2.3–5.1 | 6.6–10.3 | 3.8 | 8.6 | 0.8 | 1.0 | 3.8 | 8.0 |
| Macroplacoid row | 30 | 13.5–30.9 | 44.4–61.8 | 23.6 | 52.9 | 4.4 | 4.6 | 25.0 | 52.5 |
| Placoid row | 28 | 17.2–37.8 | 53.6–75.6 | 28.9 | 65.3 | 5.4 | 5.2 | 30.2 | 63.4 |
| Claw 1 lengths | |||||||||
| External primary branch | 22 | 7.7–12.4 | 21.4–26.3 | 10.8 | 24.5 | 1.5 | 1.2 | 11.9 | 25.0 |
| External secondary branch | 18 | 6.0–10.3 | 16.4–21.4 | 8.3 | 18.7 | 1.2 | 1.3 | 9.5 | 20.0 |
| Internal primary branch | 22 | 7.3–11.5 | 20.3–24.9 | 10.1 | 23.0 | 1.4 | 1.2 | 11.0 | 23.1 |
| Internal secondary branch | 18 | 5.8–9.1 | 15.3–18.9 | 7.8 | 17.3 | 1.0 | 1.0 | 8.2 | 17.2 |
| Claw 2 lengths | |||||||||
| External primary branch | 26 | 7.9–13.4 | 22.5–28.3 | 11.4 | 26.1 | 1.8 | 1.5 | 13.2 | 27.7 |
| External secondary branch | 24 | 6.1–10.3 | 16.2–21.6 | 8.5 | 19.6 | 1.4 | 1.2 | 10.3 | 21.6 |
| Internal primary branch | 25 | 7.3–12.5 | 20.7–26.7 | 10.6 | 24.2 | 1.7 | 1.4 | 11.4 | 23.9 |
| Internal secondary branch | 22 | 5.4–9.7 | 16.0–20.5 | 8.0 | 18.5 | 1.3 | 1.1 | 9.4 | 19.7 |
| Claw 3 lengths | |||||||||
| External primary branch | 27 | 8.2–13.8 | 22.7–29.1 | 11.7 | 26.2 | 1.7 | 1.5 | 13.0 | 27.3 |
| External secondary branch | 27 | 6.5–11.1 | 16.2–23.3 | 8.9 | 19.9 | 1.3 | 1.5 | 11.1 | 23.3 |
| Internal primary branch | 25 | 8.0–13.0 | 22.8–27.4 | 10.9 | 24.5 | 1.5 | 1.2 | 12.5 | 26.3 |
| Internal secondary branch | 21 | 5.5–10.1 | 16.3–21.3 | 8.4 | 18.8 | 1.3 | 1.4 | 9.8 | 20.6 |
| Claw 4 lengths | |||||||||
| Anterior primary branch | 23 | 9.0–14.9 | 21.8–31.0 | 12.1 | 27.2 | 1.7 | 1.8 | 12.0 | 25.2 |
| Anterior secondary branch | 20 | 6.5–11.1 | 17.9–23.4 | 8.8 | 19.9 | 1.3 | 1.2 | 8.8 | 18.5 |
| Posterior primary branch | 22 | 9.1–14.8 | 24.8–31.6 | 12.7 | 28.7 | 1.7 | 1.5 | 13.5 | 28.4 |
| Posterior secondary branch | 21 | 6.4–11.2 | 17.9–23.6 | 9.3 | 21.1 | 1.4 | 1.6 | 10.4 | 21.8 |
Notes:
- N
-
number of specimens/structures measured, RANGE refers to the smallest and the largest structure among all measured specimens
- SD
-
standard deviation
- pt
-
ratio of the length of a given structure to the length of the buccal tube expressed as a percentage
Type locality: 36°18′N, 82°22′W, 517 m asl; north side of East Tennessee State University campus, Johnson City, Washington County, TN, USA, moss (Grimmia sp.) on concrete caps of brick fence posts; xerothermic habitat fully exposed to sun and wind.
Etymology: This species is named for the shape of the projections on the eggs, which resemble the famous chocolate candy, Hershey’s Kisses. From basiatus (L.), which means “kissed”
Type material: Holotype and 234 paratypes (199 specimens, 35 eggs –2 embryonated).
Type depositories: The holotype and 9 paratypes are deposited in the National Museum of Natural History, Smithsonian Institution, Washington, DC, USA (holotype slide number USNM 1622560; paratype slide numbers USNM 1622561, USNM 1622562, USNM 1622563, USNM 1622564, USNM 1622565 (2 specimens), USNM 1622566, USNM 1622567, USNM 1622568). Additional paratypes are deposited in the Nelson Collection (180 slides), East Tennessee State University, Johnson City, TN, USA; Bertolani Collection (12 slides with 20 animals and 2 eggs), University of Modena and Reggio Emilia, Modena, Italy; and Kaczmarek Collection (34 slides), Adam Mickiewicz University, Poznań, Poland.
| CHARACTER | N | RANGE (μ m) | MEAN (μ m) | SD (μ m) |
|---|---|---|---|---|
| Diameter of egg without processes | 8 | 72.3–88.9 | 82.2 | 6.6 |
| Diameter of egg with processes | 8 | 96.0–113.3 | 104.6 | 5.9 |
| Process height | 25 | 10.9–17.0 | 13.8 | 1.5 |
| Process base width | 25 | 14.2–19.0 | 16.4 | 1.2 |
| Process base/height ratio | 25 | 101%–142% | 120% | 12% |
| Distance between processes | 23 | 3.9–7.5 | 5.3 | 0.9 |
| Number of processes on the egg circumference | 8 | 14–14 | 14.0 | 0.0 |
Notes:
Description of the new species
Animals (measurements and statistics in Table 4)
Body colour white (transparent after fixation in Hoyer’s medium; Figs. 6A–6B). Larger specimens with several orange or brown irregular excretory granules. Eyes present in 97% of measured specimens; eyes were absent only in one specimen. Entire cuticle covered with conspicuous small, round pores (1.0–1.5 in diameter) distributed randomly but more clearly visible on the anterior part of the body (Figs. 6C–6E). Clearly visible granulation present on all legs, but much more indistinct on legs I-III (Figs. 7A–7C). Buccal-pharyngeal apparatus of the Macrobiotus type, with the ventral lamina and ten peribuccal lamellae (Figs. 8A–8B). Mouth antero-ventral. The oral cavity armature of the hufelandi type (Kaczmarek & Michalczyk, 2017), with three bands of teeth visible under PCM (Figs. 8C–8D). The first band of teeth composed of very small granules (PCM), arranged in a few rows and situated on the basal parts of the lamellae and just below them. The second band of teeth composed of small granules (PCM), arranged in a few rows, just above the third band of teeth. The third band of teeth composed of three dorsal and three ventral teeth in the shape of transverse ridges (PCM). Median tooth on ventral side in shape of an oval granule that can sometimes be divided into two parts. Pharyngeal bulb spherical with triangular apophyses, two rod-shaped macroplacoids and a thin microplacoid. Macroplacoid length configuration 2<1. The first macroplacoid with a central constriction (Fig. 8E). The second macroplacoid without constriction, but with latero-terminal globular projections. Claws of the hufelandi type, stout (Figs. 9A–9D). Primary branches with distinct accessory points. Lunules I–III smooth, but IV slightly indented, with very tiny teeth mainly visible with SEM (Figs. 9C insert and 9D). Thin paired bars under claws I–III present. Other cuticular thickenings on legs absent.
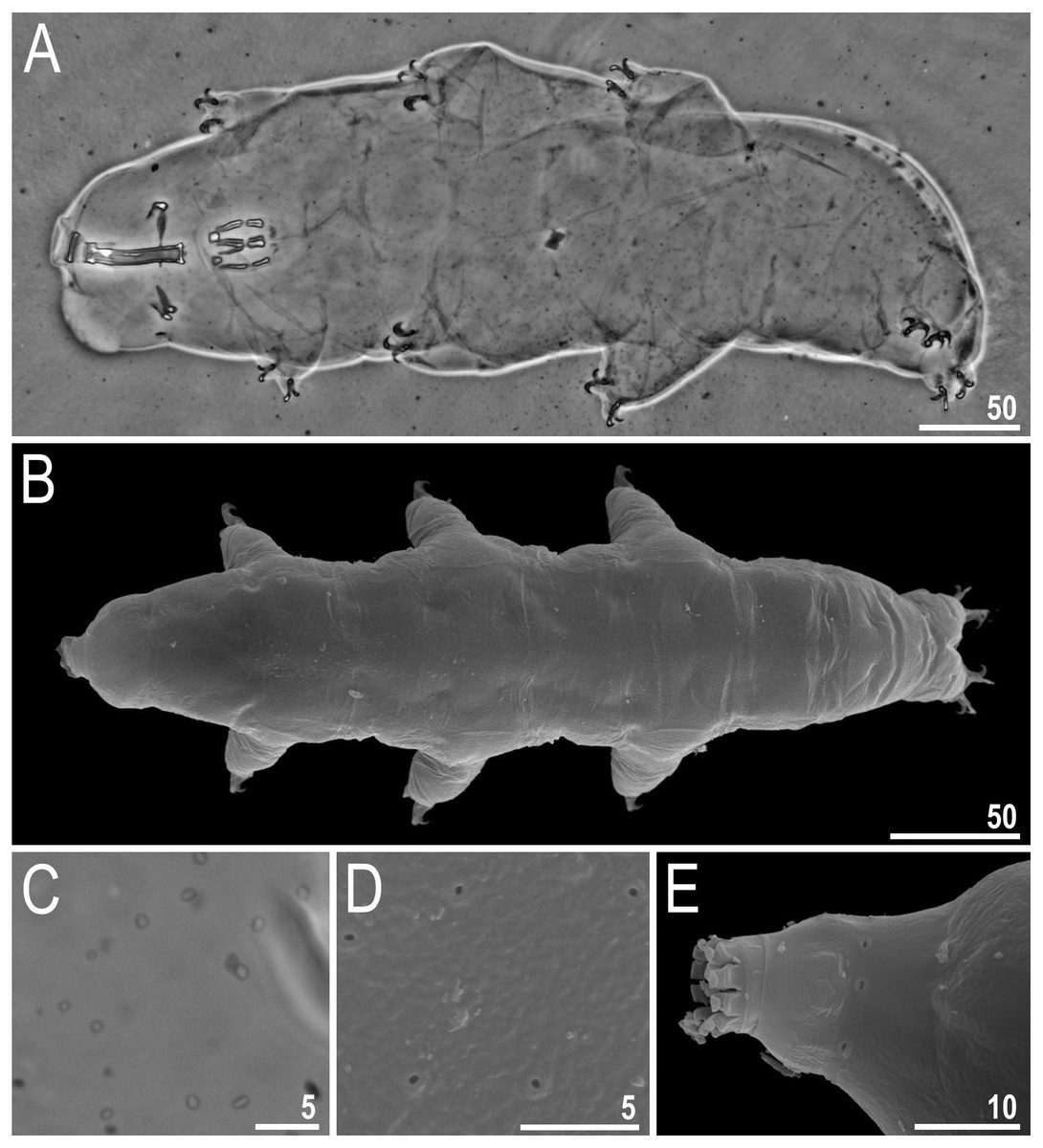
Figure 6: Macrobiotus basiatus sp. nov.
(A) Habitus, dorsoventral assembled image (holotype, PCM). (B) Animal in dorsal view (paratype, SEM). (C) Cuticle covered with pores (holotype, PCM). (D) Cuticle covered with pores (paratype, SEM). (E) Ring of pores visible posterior to the mouth opening. Scale bars in micrometres [µm]. Photo credit: Milena Roszkowska.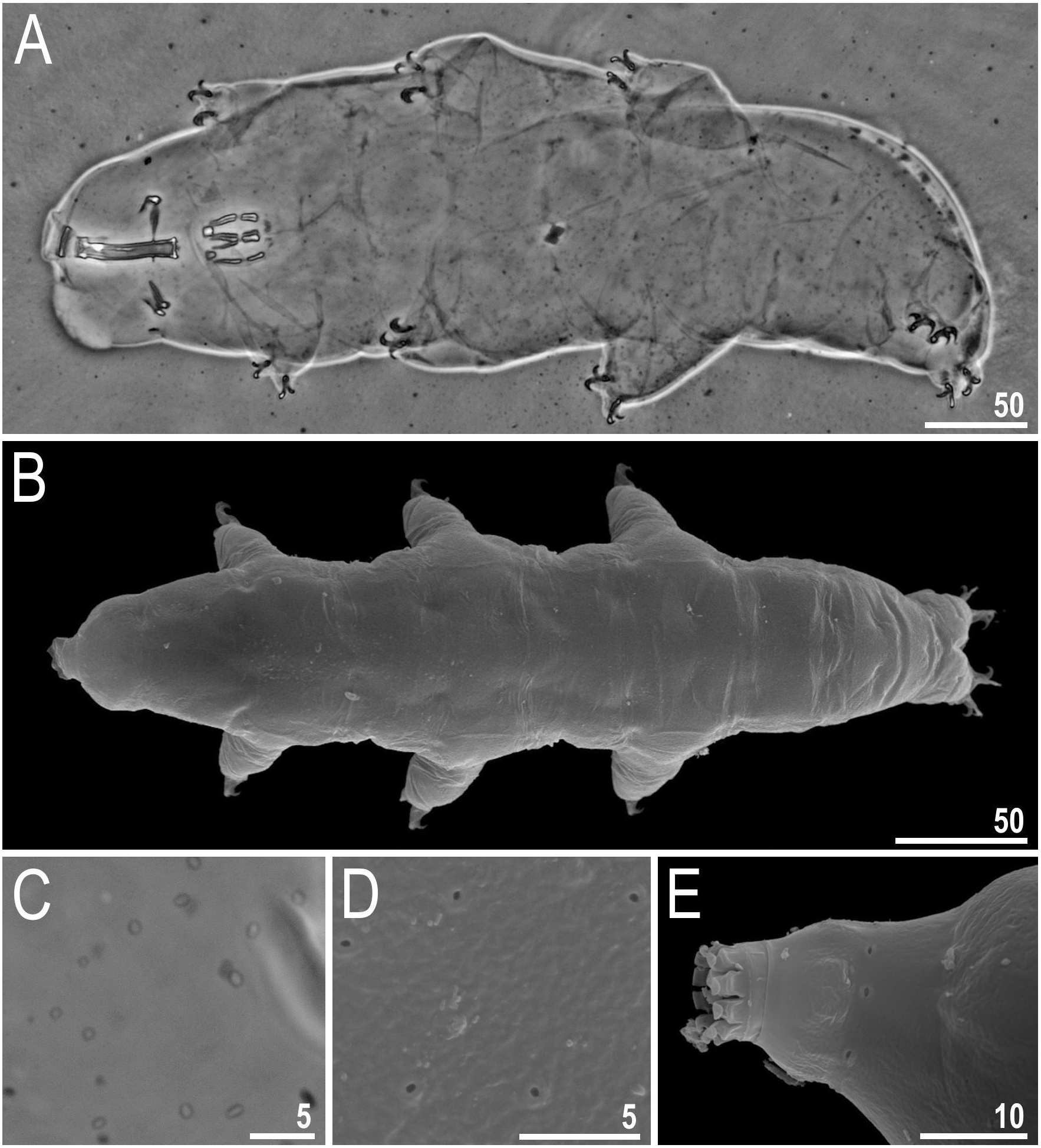{kind=link}
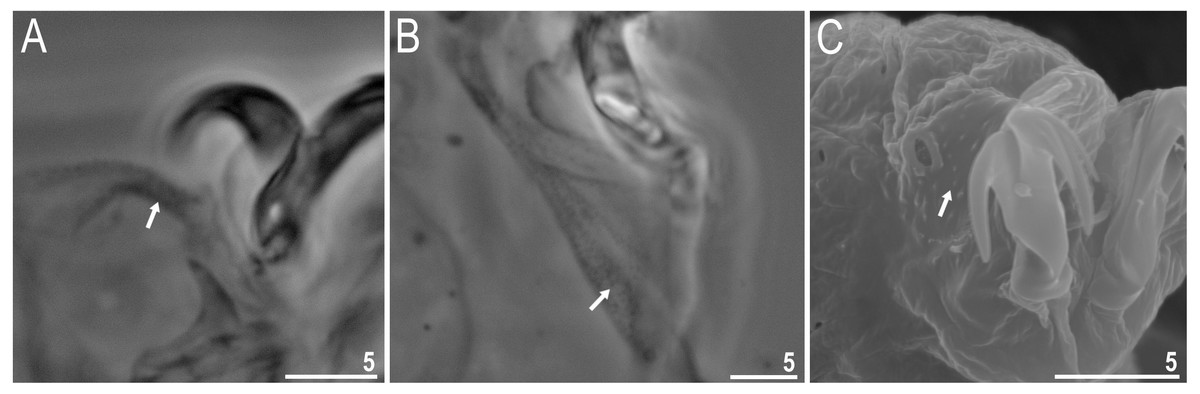
Figure 7: Macrobiotus basiatus sp. nov. granulation on legs (filled arrows).
(A) Leg III (paratype, PCM). (B) Leg IV (paratype, PCM). (C) Leg IV (paratype, SEM). Scale bars in micrometres [µm]. Photo credit: Milena Roszkowska.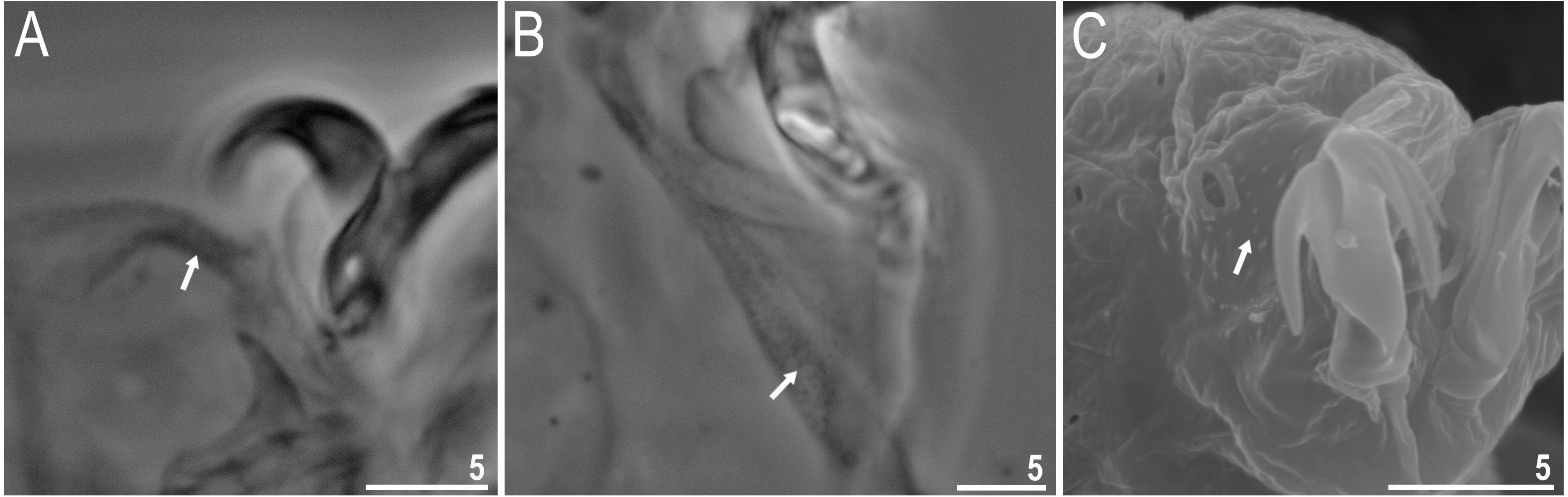{kind=link}
Eggs (measurements and statistics in Table 5)
Eggs spherical, ornamented and laid freely (Figs. 10A–10B). Processes conical with short and sometimes a slightly flexible elongated tip (Figs. 10A–10F). In PCM, the surface of the processes and arms of the areoles have a reticular pattern with a regular rounded mesh (Figs. 10C–10D). In SEM, the processes and arms of areoles are smooth and visibly raised above the internal surface of the egg and connected with it by branched columns (Figs. 10E– 10F). Each process is surrounded by 9–10 areoles, smooth inside (Figs. 10D–10H), not always visible in PCM (Fig. 10B). The number of areoles between two neighboring processes is always two (Fig. 10H).
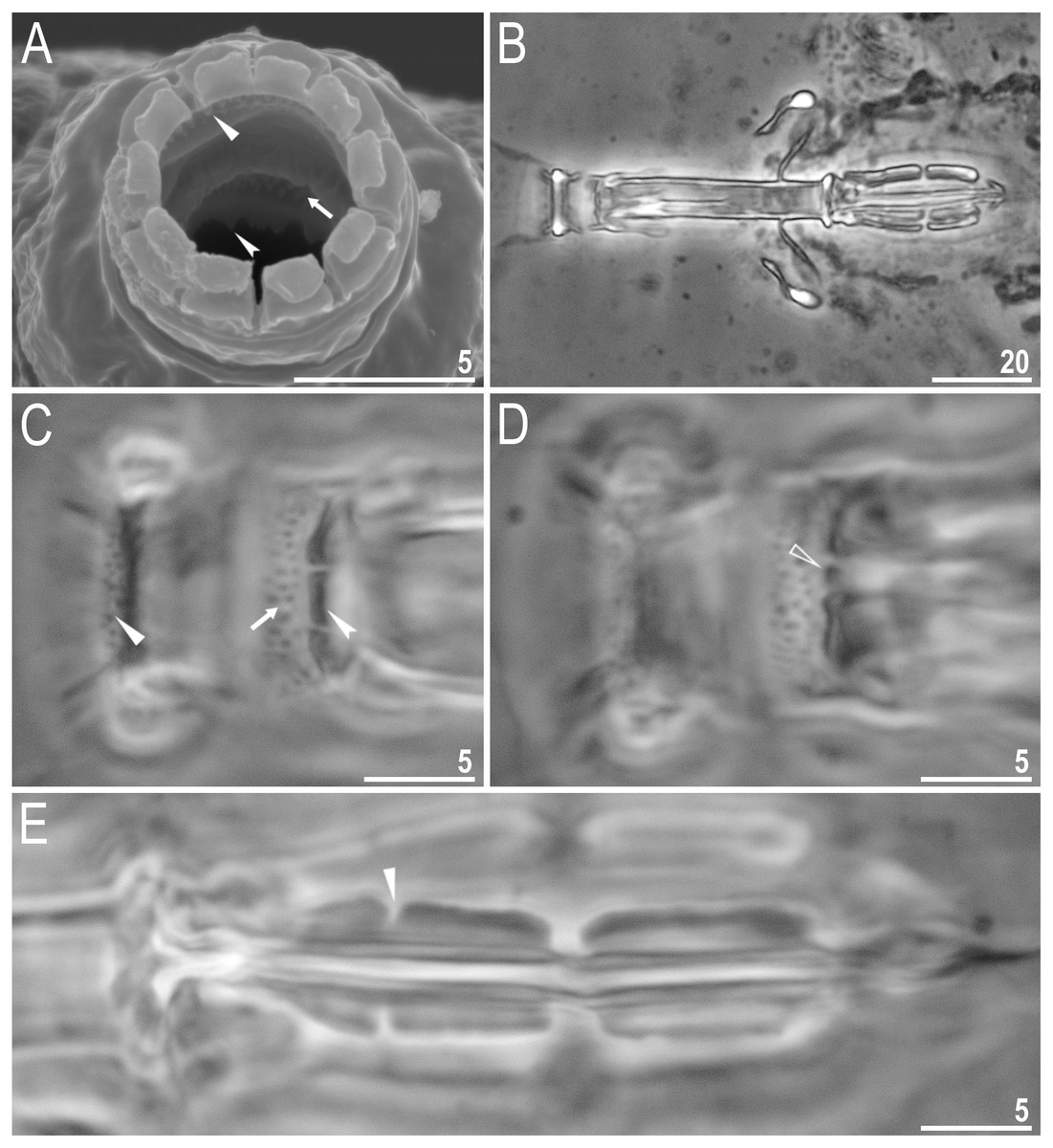
Figure 8: Macrobiotus basiatus sp. nov. claws.
(A) Mouth opening with ten peribuccal lamellae, filled arrowhead indicates teeth of the first band, filled arrow indicates teeth of the second band and indented filled arrowhead indicates teeth of the third band (paratype, SEM). (B) Buccalpharyngeal apparatus (dorsoventral assembled image) (paratype, PCM). (C) Oral cavity armature (dorsal view), filled arrowhead indicates teeth of the first band, filled arrow indicates teeth of the second band and indented filled arrowhead indicates teeth of the third band. (D) Oral cavity armature (ventral view), empty arrowhead indicates median tooth of the third band on ventral side in shape of oval granule (paratype, PCM). (E) Macroplacoids, filled arrowhead indicates the first macroplacoid with a central constriction (paratype, PCM). Scale bars in micrometres [µm]. Photo credit: Milena Roszkowska.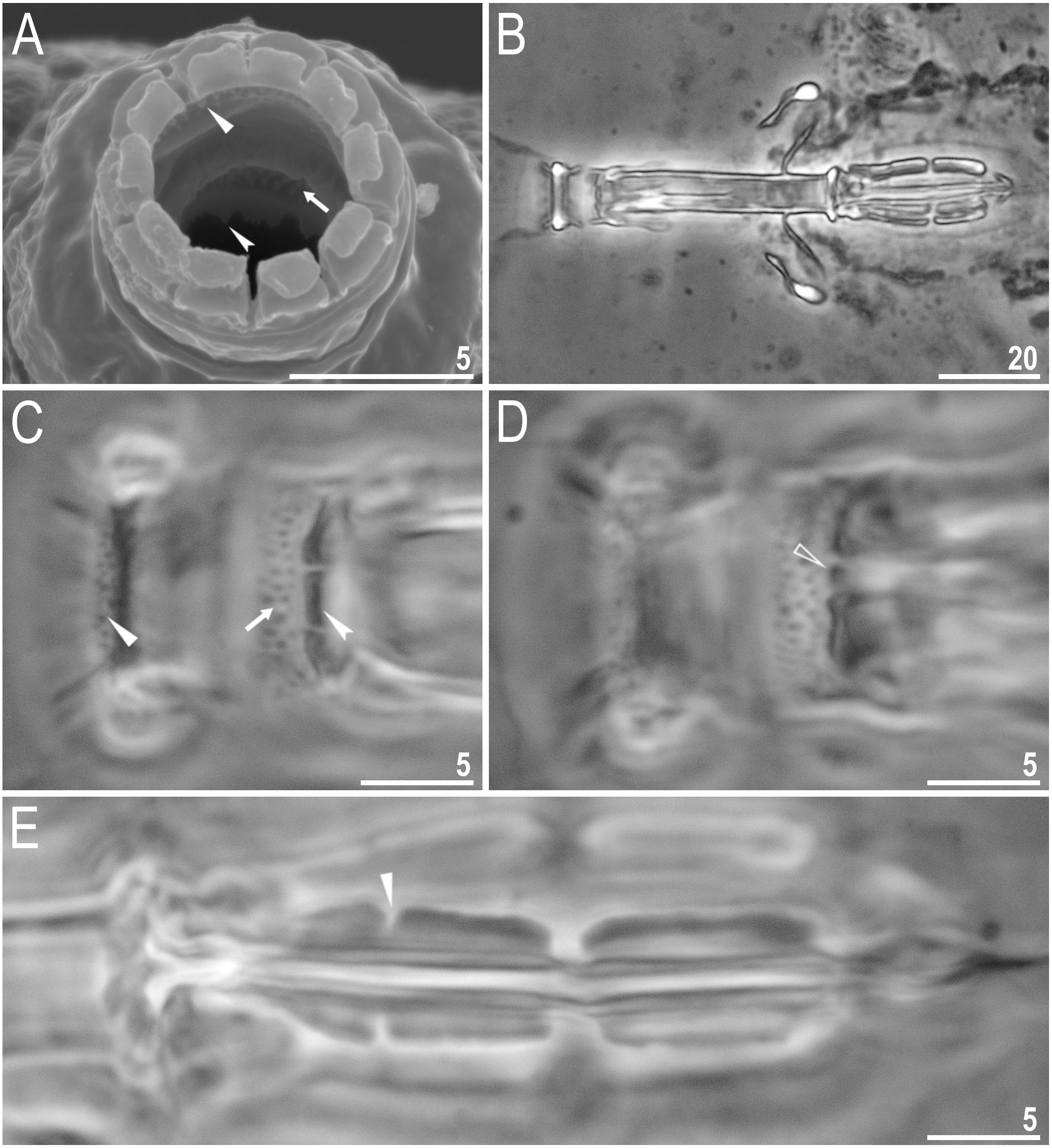{kind=link}
Differential diagnosis
The new species, based on the morphology of the eggs and adults, is most similar to Macrobiotus deceptor Meyer et al., 2017, Macrobiotus nelsonae Guidetti, 1998, Macrobiotus pallarii Maucci, 1954, and Macrobiotus ragonesei Binda et al., 2001, but it differs specifically from:
-
Macrobiotus deceptor, known only from Saint Mary Parish, Antigua (Meyer et al., 2017), by: presence of eyes, cuticular pores, and granulation on legs in the new species, oval areoles around egg processes (polygonal in Mac. deceptor), shorter egg processes (10.9–17.0 µm in the new species vs 21.2–30.3 µm in Mac. deceptor) and smaller number of areoles around egg processes (ca. 8 in the new species vs 10–12 in Mac. deceptor).
-
Macrobiotus nelsonae, known only from Roan Mountain, Tennessee, USA (Guidetti, 1998), by: the stylet supports situated in more anterior position (pt 74.4–78.9 in new species vs 79.4–84.5 in Mac. nelsonae), shorter egg processes (10.9–17.0 µm in new species vs 20.8–31.7 µm in Mac. nelsonae), oval areoles around egg processes in the new species (polygonal in Mac. nelsonae), narrower egg processes (14.2–19.0 µm in new species vs 20.0–34.7 µm in Mac. nelsonae) and smaller number of areoles around egg processes (ca. 8 in new species vs 11–12 in Mac. nelsonae).
-
Macrobiotus pallarii, reported from Asia, Europe and North America (McInnes, 1994), by: the presence of slightly indented with very tiny teeth on lunules (visible with SEM) on legs IV (smooth lunules in Macrobiotus pallarii) and presence of two rows of areoles around each egg processes (only one row of areoles in Mac. pallarii). Macrobiotus pallarii, is poorly described and needs revision, but we have included it here for comparison with the new species.
-
Macrobiotus ragonesei, reported from North Kivu Province, Democratic Republic of the Congo (Binda et al., 2001), by: the presence of granulation on legs I–III, the presence of regular reticular mesh on the entire surface of the egg processes (egg processes with only the basal part covered with regular reticular mesh in Mac. ragonesei), higher number of processes on the egg circumference (14 in the new species vs 10–12 in Mac. ragonesei), smaller eggs (72.3–88.9 µm and 96.0–113.3 µm with and without processes, respectively, in the new species vs 92.0–100.0 µm and 126.0–136.0 µm with and without processes, respectively, in Mac. ragonesei), shorter egg processes (10.9–17.0 µm in the new species vs 20.0–23.0 µm in Mac. ragonesei), and narrower egg processes (14.2–19.0 µm in the new species vs 20.0–26.0 µm in Mac. ragonesei).
DNA sequences
Good quality sequences described here are accessible via GenBank accession numbers MT488397, MT488398, MT488399, MT488400, MT488401, MT488402 for 28S rRNA, MT498094 for 18S rRNA, MT505165, MT505166, MT505167, MT505168, MT505169 and MT505170 for ITS-2, and MT502116 for COI. We obtained two exoskeletons of Mac. basiatus sp. nov. after DNA isolation, which are deposited in the Kaczmarek collection, labelled USA/NEL/4/S and USA/NEL/6/S.
Molecular differential diagnosis
All the obtained DNA barcode sequences of Mac. basiatus sp. nov. were unique and distinct from those deposited in GenBank.
The p-distances between new species and other deposited in GenBank are:
-
18S rRNA: 0.4–3.6% (1.9% on average), with the most similar being the Mac. hufelandi group (FJ435738 and FJ435739) (Guil & Giribet, 2012) and the least similar being Macrobiotus polypiformis Roszkowska et al., 2017 (KX810008) (Roszkowska et al., 2017).
-
28S rRNA: 1.8–11.8% (6.6% on average), with the most similar being Mac. hufelandi group (FJ435751 and FJ435754 –FJ435755) (Guil & Giribet, 2012) and the least similar being Mac. polypiformis (KX810009) (Roszkowska et al., 2017).
-
COI: 15.3–23.4% (19.1% on average), with the most similar being Macrobiotus terminalis Bertolani & Rebecchi, 1993 (AY598775) (Guidetti et al., 2005) and the least similar being Mac. polypiformis (KX810012) (Roszkowska et al., 2017).
-
ITS-2: 0.5–30.6% (17.2% on average), with the most similar being Macrobiotus canaricus (Stec, Krzywański & Michalczyk, 2018) (MH063929) (Stec, Krzywański & Michalczyk, 2018) and the least similar being Macrobiotus scoticus Stec et al., 2017 (KY797268) (Stec et al., 2017).
Discussion
The most numerous species, Vir. viridissimus, was obviously a well-established species on the fence posts. With the second highest relative abundance, Mil. inceptum was also considered highly successful in this xerothermic habitat. Previously found in samples from this site (Nelson & Adkins, 2001) but undescribed until now, Mac. basiatus sp. nov. was relatively low in abundance.
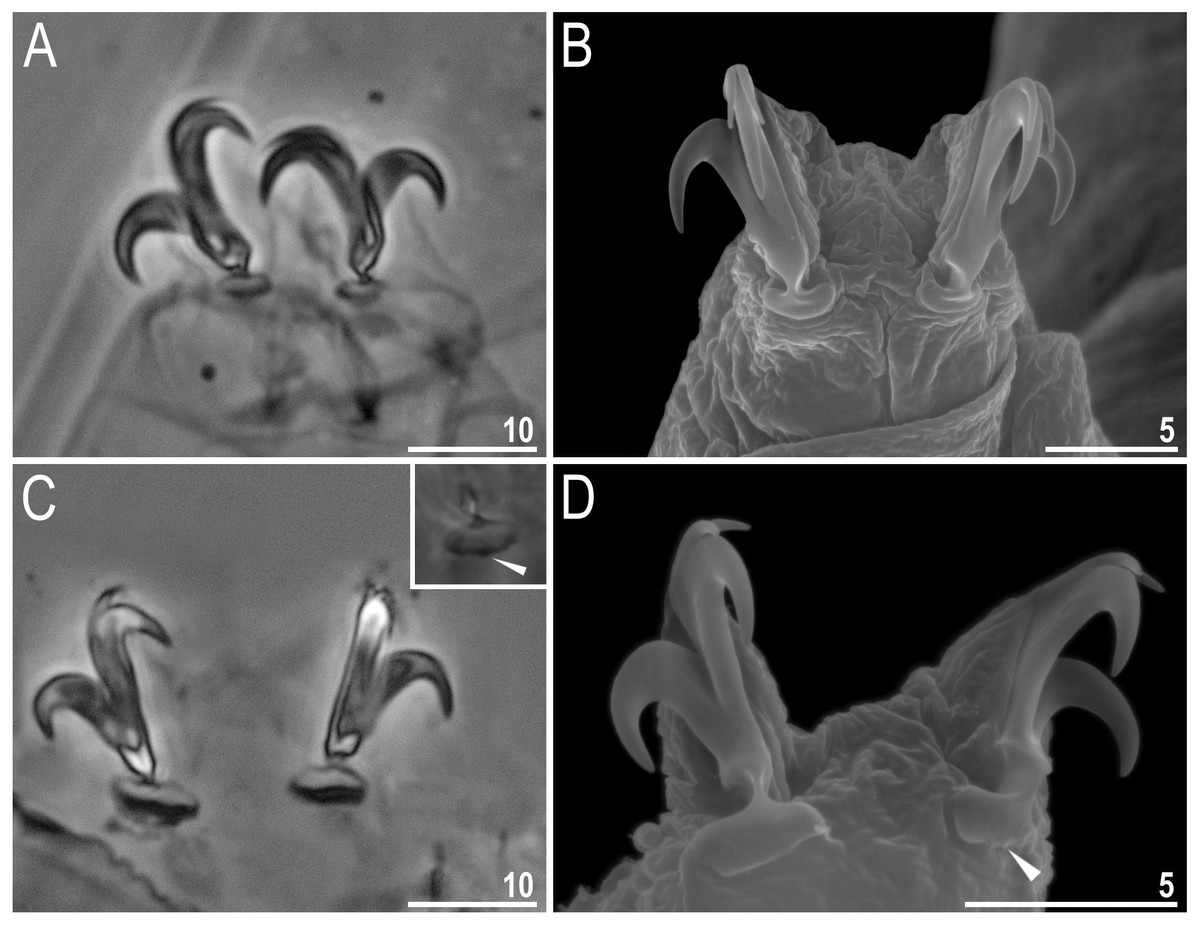
Figure 9: Macrobiotus basiatus sp. nov. claws.
(A) Claws III with smooth lunules (paratype, PCM). (B) Claws II with smooth lunules (paratype, SEM). (C) Claws IV with smooth lunules, insert shows barely visible teeth found on one specimen (filled arrowhead) (paratype, PCM). (D) Claws IV with lunules with very tiny teeth visible (filled arrowhead) (paratype, SEM). Scale bars in micrometres [µm]. Photo credit: Milena Roszkowska.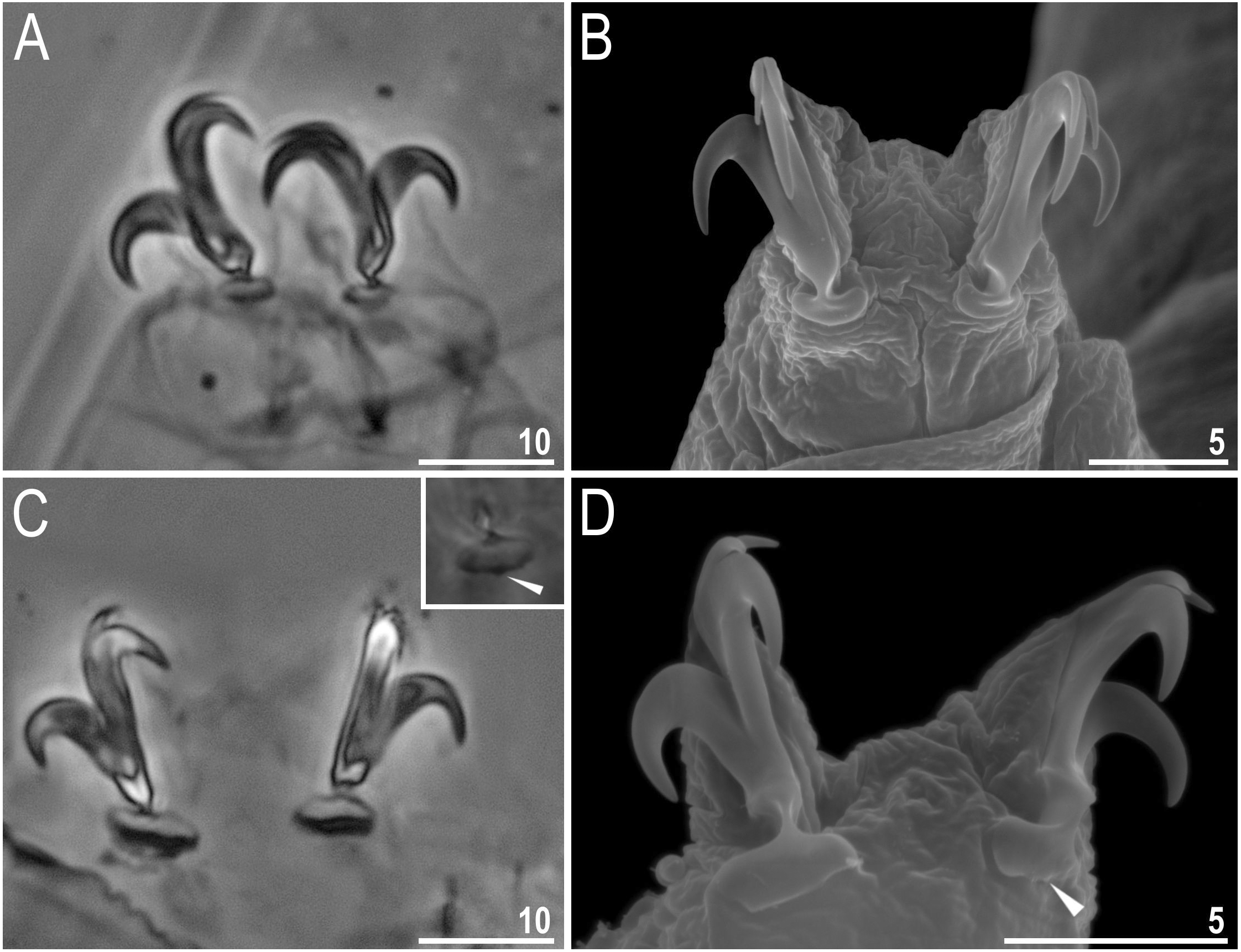{kind=link}

Figure 10: Macrobiotus basiatus sp. nov. eggs.
(A) Entire egg (SEM). (B) Entire egg (PCM). (C–D) Egg processes, indented arrowheads indicate visible arms of areoles (PCM). (E–F) Egg processes, indented arrowheads indicate branched columns under processes and arms of areoles (SEM). (G) Egg surface with areoles (PCM). (H) Egg surface with areoles (SEM). Scale bars in micrometres [µm]. Photo credit: Milena Roszkowska.{kind=link}
The large number of Mil. inceptum individuals is interesting since this species is a predator. If Milnesium individuals fed on Viridiscus or Macrobiotus specimens, then cuticular parts of the animals (e.g., claws, buccal tubes, dorsal plates) should have been visible within the gut, however, no evidence of this was found. Thus, the major food supply for such a large population of this predator is probably constituted by the numerous rotifers and nematodes, verified by the presence of rotifer jaws in guts of some Mil. inceptum specimens.
The genus Viridiscus was recently established by Gąsiorek & Michalczyk in Gąsiorek et al. (2019) for the viridis group of species previously included in the genus Echiniscus. Viridiscus is characterized by species with a fully or partially green cuticle (although sometimes brown, gray, or almost black), a dorsal plate sculpture composed of an intracuticular foam-like layer and small, flat and densely arranged epicuticular granules, no cuticular appendages other than cephalic appendages and lateral cirri A, well-developed sabre-like claws, absence of males, and inhabiting terrestrial ecosystems. The genus includes Vir. clavispinosus, Vir. perviridis, Vir. rufoviridis, Vir. viridis, Vir. viridianus, Vir. viridissimus, and now Vir. miraviridis sp. nov. The dorsal cuticle is characterized by an evident surface (with “pits”, granules, meshes, etc.) characteristic of each species, overlying a quite uniform background of fine dots. In Viridiscus a ventral cuticular pattern is also present, but it is also present in species from different heterotardigrade genera; this character has not yet been evaluated for its taxonomic value in the genus. The presence of a green cuticle, a primary morphological criterion for Viridiscus, is also found in Echiniscus pooensis Rodriguez-Roda, 1948, which has lateral and dorsal spines on the cuticular plates and therefore was not included in the genus in Gąsiorek et al. (2019).
We noted also that the specimens of Vir. rufoviridis of the Maucci collection present a different structure of the dorsal cuticle of the paired plates and second median plate than any other species of Viridiscus, which was also reported by Pilato, Fontoura & Lisi (2008). In fact, the granules present in the anterior position of these plates are not overlying the background of fine dots, but these granules become smaller (around the middle portion of each plate) forming the background of small dots present under the posterior surface of the plates characterized by white “pits.”
Males are very rare in the genera derived from the evolutionary line of the former Echiniscus “arctomys group” according to Gąsiorek et al. (2019). Only 37 years ago, males were considered completely absent in the widespread genus Echiniscus, at that time composed of about 112 species (Ramazzotti & Maucci, 1983), including the former viridis group (now the genus Viridiscus) within the “arctomys group”. The first report of males in Echiniscus species was published by Dastych (1987) and later by others (e.g., Claxton, 1996; Miller, Claxton & Heatwole, 1999; Mitchell & Romano III, 2007; Claxton & Dastych, 2017, who reported males in several species of Echiniscus. Miller, Claxton & Heatwole (1999) hypothesized a Gondwana origin of bisexual Echiniscus species, comprising those of the “arctomys group.” Although Mitchell & Romano III (2007) reported the first males in the former Echiniscus mauccii [now Claxtonia mauccii (Ramazzotti, 1956)] in North America, our finding of males in Vir. miraviridis sp. nov. confirms the presence of males in the genus Viridiscus on the continent. Therefore, the proposal of a Gondwana origin of bisexual species in this evolutionary line needs to be re-evaluated. In the description of the new genus Viridiscus in Gąsiorek et al. (2019), the species in the genus were reported to reproduce by parthenogenesis. However, in our study, males of Vir. viridissimus were rare (0.04%) but present, raising questions about that one aspect of the generic description. Additional observations of gonopores may reveal rare males in other species of Viridiscus.
The distribution of the genus Viridiscus is global, occurring throughout Europe, North and South America and on Pacific and Atlantic islands, leading to the hypothesis that the genus is also present in areas not yet well investigated. Although Vir. viridis has been widely reported from Europe, Hawaii (Oahu Is.) USA, North and South America, at present only the Hawaiian reports are correct according to Pilato, Fontoura & Lisi (2008). A similar wide range (Europe, North and South America) has been reported for Vir. viridissimus, but probably this species also has a narrower distribution. Viridiscus perviridis is reported from Europe and North America, Vir. viridianus from North America and the Azores Islands (Atlantic Ocean), and Vir. rufoviridis only from South America. Viridiscus clavispinosus is currently considered endemic to Santo Antão Island (Republic of Cape Verde, Atlantic Ocean, off West Africa) and Viridiscus miraviridis sp. nov. is known only from Tennessee (USA, North America) (McInnes, 1994; Fontoura, Pilato & Lisi, 2011; Kaczmarek, Michalczyk & McInnes, 2015; Kaczmarek, Michalczyk & McInnes, 2016). The records of the widely distributed species need further confirmation.
Conclusions
Five species of tardigrades, including two species new to science, were found in moss from a xerothermic habitat in Tennessee: Milnesium inceptum (Eutardigrada), Macrobiotus basiatus sp. nov. (Eutardigrada), and three species of Heterotardigrada, Viridiscus. miraviridis sp. nov., Vir. perviridis, and Vir. viridissimus. This is the first record of Mil. inceptum from North America, confirmed by COI analysis, and the first record of males in the genus Viridiscus. The two new species were identified by an integrative analysis of morphological and/or molecular characters. This research increases our knowledge of tardigrades in Tennessee and North America and adds to the database on biogeography and distribution. Further investigations could examine the tardigrade community structure, analysing the co-existence of multiple Viridiscus species in the isolated habitats, as well as determining the chemical composition of the green pigment in the Viridiscus cuticle, as little is known of the chemical composition of various pigments in tardigrades.