
New genicular joint angle criteria for flexor muscle (Musculus Semimembranosus) during the terrestrial mammals walking
- Published
- Accepted
- Received
- Academic Editor
- Virginia Abdala
- Subject Areas
- Animal Behavior, Paleontology, Zoology
- Keywords
- Knee joint angle, Muscle running, Mammalian walk, Co-contraction, Positional relationships of bones
- Copyright
- © 2023 Mizuno and Kohno
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2023. New genicular joint angle criteria for flexor muscle (Musculus Semimembranosus) during the terrestrial mammals walking. PeerJ 11:e15379 https://doi.org/10.7717/peerj.15379
Abstract
Background
The genicular or knee joint angles of terrestrial mammals remain constant during the stance phase of walking; however, the angles differ among taxa. The knee joint angle is known to correlate with taxa and body mass among extant mammals, yet several extinct mammals, such as desmostylians, do not have closely related descendants. Furthermore, fossils lose their soft tissues by the time they are unearthed, making body mass estimates difficult. These factors cause significant problems when reconstructing the proper postures of extinct mammals. Terrestrial mammals use potential and kinetic energy for locomotion; particularly, an inverted pendulum mechanism is used for walking. This mechanism requires maintaining the rod length constant, therefore, terrestrial mammals maintain their joint angle in a small range. A muscle reaction referred to as co-contraction is known to increase joint stiffness; both the agonist and antagonist muscles work simultaneously on the same joint at the same time. The musculus semimembranosus flexes the knee joint and acts as an antagonist to muscles that extend it.
Methods
Twenty-one species of terrestrial mammals were examined to identify the elements that constitute the angle between the m. semimembranosus and the tibia based on the period between the hindlimb touching down and taking off from the ground. Measurements were captured from videos in high-speed mode (420 fps), selecting 13 pictures from the first 75% of each video while the animals were walking. The angles between the main force line of the m. semimembranosus and the tibia, which were defined as θsm−t, were measured.
Results
The maximum and minimum angles between the m. semimembranosus and the tibia (θsm−t) of the stance instance (SI) were successfully determined for more than 80% of the target animals (17 out of 21 species) during SI-1 to SI-13 within ±10° from the mean. The difference between each successive SI was small and, therefore, the θsm−t transition was smooth. According to the results of the total stance differences among the target animals, θsm−t was relatively constant during a stance and, therefore, average θsm−t (θave) can represent each animal. Only Carnivora had a significant difference in the correlation between body mass and θave. In addition, there were significant differences in θave between plantigrade and unguligrade locomotion.
Conclusion
Our measurements show that θave was 100 ± 10° regardless taxon, body mass, and locomotor mode. Thus, only three points on skeletons need to be measured to determine θave. This offers a new approximation approach for understanding hindlimb posture that could be applied to the study of the hindlimbs of extinct mammals with no closely related extant descendants.
Introduction
Hindlimbs act as propulsive devices for terrestrial locomotion (Demes et al., 1994). Common terrestrial behaviors require limbs to support body mass against gravity, which means that terrestrial mammals must resist collapsing joints against gravity, requiring the maintenance of extending joints. Although limbs have the same role in that supporting body mass, joint angles differ between species (Biewener, 1983, 2005; Inuzuka, 1996; Dutto et al., 2006; Polly, 2007; Fujiwara, 2009; Dick & Clemente, 2017). For example, the angles at the knee joint in Asian elephants is approximately 160° (Ren et al., 2008), compared to 137° in chacma baboons (Patel et al., 2013), 115° in domestic cats, 124° in lions (Day & Jayne, 2007). Thus, the limb joint angle is unique to each species; however, the joints have a wider rotatable range than the angle maintained during standing or walking. This causes problems when reconstructing skeletal specimens into an accurate posture when they were alive. In particular, extinct taxa present a significant challenge when reconstructing postures because the angle when they were alive cannot be observed. For example, desmostylian mammals, which do not have closely related living descendants, have been reconstructed in several different postures even though almost complete skeletons of the same species have been unearthed (Domning, 2002; Inuzuka, Sawamura & Watabe, 2006; Fujiwara, 2009). Furthermore, earlier diverging cetaceans, such as pakicetids and ambulocetids, had functional hindlimbs, and extant cetaceans had completely lost their hindlimbs (Thewissen, Madar & Hussain, 1998; Gingerich, 2001; Thewissen et al., 2001; Madar, 2007; Gingerich et al., 2009, 2017). In such cases, there are no extant mammals that can be used as references for skeletal reconstruction. Therefore, knowledge of hindlimb postures in terrestrial mammals is important to understand the transition of locomotive ability through mammalian evolution, including the adaptation from land to sea.
Several studies have explored the relationship between limb posture and variables such as taxa, body mass, and skeletal morphology in extant mammals (Biewener, 1983, 1989, 1990, 2005; Day & Jayne, 2007; Fujiwara, 2009; Fujiwara & Hutchinson, 2012: Dick & Clemente, 2017). These studies indicate that the larger the size of the mammal species the more upright limb posture the species has. However, there are several exceptions to the relationship between limb posture and body mass (Fujiwara, 2009). Furthermore, there is a significant problem with estimating the body mass of extinct mammals because fossils have already lost soft tissues by the time they are unearthed. To resolve these problems, it is important to identify joint angle criteria that are unaffected by other factors as possible.
Quadrupedal mammals use potential and kinetic energy to accelerate their center of mass during running and walking (Cavagna, Heglund & Taylor, 1977; Alexander & Jayes, 1978; Hildebrand, 1984; Hildebrand & Hurley, 1985; Alexander, 1991; Griffin, Main & Farley, 2004), employing an inverted pendulum movement to walk. This movement allows the quadrupedal mammals to generate the necessary energy to lift and accelerate the center of mass and maintain a constant stride length (Cavagna, Heglund & Taylor, 1977; Griffin, Main & Farley, 2004). The inverted pendulum requires that the distance between the ground and the center of mass is constant; therefore, the limb joints are maintained within limited range while walking (Manter, 1938; Gray, 1944; Goslow, Reinking & Stuart, 1973; Goslow et al., 1981; Alexander & Jayes, 1983; Inuzuka, 1996; Fischer et al., 2002; McGowan, Baudinette & Biewener, 2005). When a joint angle is locked against the force of gravity, not only the agonist muscle but also the antagonist muscle work together. This action increases joint stiffness in humans (Olmstead et al., 1986; Louie & Mote, 1987; Nielsen et al., 1994; Riemann & Lephart, 2002; Knarr, Zeni & Higginson, 2012). Some electromyographic studies of quadrupedal mammals have shown that both agonist and antagonist muscles act simultaneously during the stance phase—the period in which the foot under consideration is in contact with the floor—when the limb supports the body mass (Engberg & Lundberg, 1969; Tokuriki, 1973; Deban, Schilling & Carrier, 2012; Araújo et al., 2016). The knee joint maintains an angle owing to extension against gravity, and the musculus semimembranosus acts as the knee joint flexor muscle, which is the antagonist muscle of the m. quadriceps femoris when the joint extends.
The m. semimembranosus attaches to the ischial tuberosity and interior proximal end of the tibia (Fig. 1) (Böhmer et al., 2020). These attachment positions do not move, and the involved parts of the skeleton do not change their shape greatly among taxa. Thus, the positional relationship between the muscle and these parts of the skeleton also shows the relationship among skeleton elements. In addition, the angles of the pelvic girdle differ among different body masses (Polly, 2007). Therefore, the angle between the line of action of the m. semimembranosus and the tibia has a smaller difference than the angle between the femur and tibia among different body masses. Here, we aimed to (1) reveal the joint angle of terrestrial mammals between the m. semimembranosus and tibia during walking, (2) explore the relationships between this angle and taxa, body mass, and locomotor mode, and (3) evaluate whether this angle might be suitable as one of the criteria for the reconstruction of hindlimb postures.
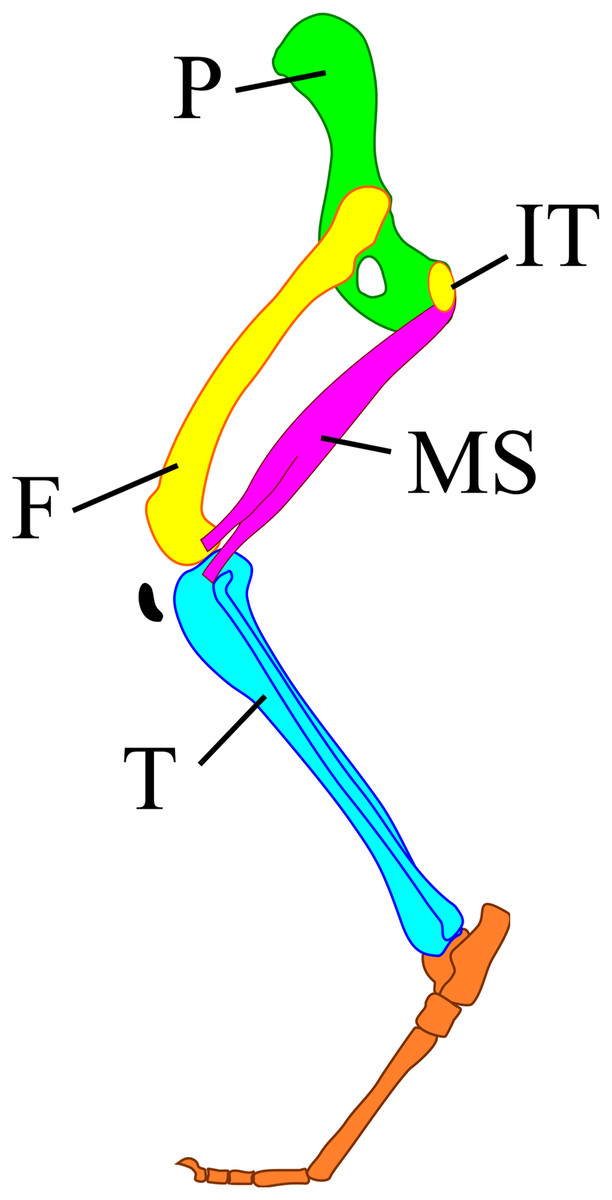
Figure 1: A model of the mammalian hindlimb bones with m. semimembranosus from the exterior view.
M. semimembranosus origin locates on the ischial tuberosity (IT) and insertion locates on the interior-proximal end of the tibia (T). The insertion of this muscle was drawn on the exterior of tibia in this figure to identify the position easily, however, the actual insertion locates on the interior of tibia. Abbreviations: F, femur; IT, ischial tuberosity; MS, m. semimembranosus; P, pelvis; T, tibia.{kind=link}
Materials and Methods
The angles between m. semimembranosus and the tibia in vivo were collected from 21 extant species from 21 genera and 14 families within seven orders (Table 1). These species were selected to cover the superorder and order of mammals (Afrotheria, Proboscidea; Euarchontoglires, Primates, Rodentia; Laurasiatheria, Artiodactyla, Carnivora, Perissodactyla; and Marsupialia, Diprotodontia), a wide range of body masses (i.e., from 0.7 kg for Suricata suricatta to 4,060 kg for Elephas maximus), and three locomotor modes (plantigrade, digitigrade, and unguligrade) (Table 1). All the target animals were kept in zoos at Higashi Park Zoological Gardens (Okazaki, Japan), Higashiyama Zoo and Botanical Garden (Aichi, Japan), Hitachi Kaminé Zoo (Ibaraki, Japan), Toyohashi Zoo and Botanical Park (Aichi, Japan), and Ueno Zoological Gardens (Tokyo, Japan), and all observations of living individuals were conducted after gaining official permissions. No significant pathologies or malformations were detected in any of the studied specimens.
| Super order | Order | Family | Genus | Species | Locomotor mode | Locality | Body mass (kg) | Body mass reference |
|---|---|---|---|---|---|---|---|---|
| Afrotheria | Proboscidea | Elephantidae | Elephas | maximus | D | UZ | 4,060.0 | Shoshani & Eisenberg (1982) |
| Euarchonto-glires | Primates | Ceropithecidae | Cercopithecus | neglectus | P | UZ | 4.5 | Fa & Purvis (1997) |
| Chlorocebus | aethiops | P | HKZ | 5.78 | Our study | |||
| Macaca | fuscata | P | HKZ | 16.0 | Obara et al. (2000) | |||
| Rodentia | Cavidae | Dolichotis | patagonum | D | HP | 8.0 | Campos, Tognelli & Ojeda (2001) | |
| Laurasia-theria | Artiodactyla | Bovidae | Ammotragus | lervia | U | UZ | 72.6 | Cassinello (1997) |
| Capra | hircus | U | HKZ | 37.0 | Our study | |||
| Cervidae | Cervus | nippon | U | UZ | 75.4 | Our study | ||
| Rangifer | tarandus | U | HZ | 120.0 | Garland (1983) | |||
| Giraffidae | Giraffa | camelopardalis | U | UZ | 1,000.0 | Garland (1983) | ||
| Carnivora | Canidae | Canis | lupus | D | HZ | 43.3 | Mech (1974) | |
| Chrysocyon | brachyurus | D | UZ | 23.0 | Our study | |||
| Felidae | Felis | catus | D | Co | 4.8 | Our study | ||
| Panthera | leo | D | TZ | 188.0 | Haas, Hayssen & Krausman (2005) | |||
| Herpestidae | Suricata | suricatta | D | HP | 0.7 | van Staaden (1994) | ||
| Ursidae | Helarctos | malayanus | P | TZ | 45.0 | Fitzgerald & Krausman (2002) | ||
| Ursus | thibetanus | P | HKZ | 100.0 | Our study | |||
| Perissodactyla | Equidae | Equus | cabullus | U | TZ | 200.0 | Garland (1983) | |
| Rhinocerotidae | Diceros | bicornis | U | UZ | 1,100.0 | Hillman-Smith & Groves (1994) | ||
| Tapiridae | Tapirus | terrestris | U | HZ | 700.0 | Padilla & Dowler (1994) | ||
| Marspialia | Diprotodontia | Macropodidae | Macropus | giganteus | O | UZ | 55.0 | Our study |
Notes:
The body mass data are the average of the data from reference or each zoo. Ambulatory style abbreviations: U, unguligrade; D, digitigrade; P, plantigrade; O, other. Institutional abbreviations: HKZ, Hitachi Kamine Zoo, Ibaraki; HP, Higashi Park Zoological Gardens, Aichi; HZ, Higashiyama Zoo and Botanical Garden, Aichi; TZ, Toyohashi Zoo and Botanical Park, Aichi; UZ, Ueno Zoo, Tokyo. Institutions are sorted by alphabetical order.
All target animals were subjected to video recording using a digital movie camera (EX-FH20; Casio, Tokyo, Japan) in high-speed mode (420 fps). The camera was mounted on a tripod along the visitor viewing route. Therefore, the distance from each target depended on the exhibition/cage arrangement. All videos were taken from the lateral side and at nearly the same level as the target animal when they walked vertically and completely (without stopping, turning, or changing speed) with the camera on flat ground. We waited until each target walked across the camera voluntarily, without any coaxing, meaning it took several weeks or months to obtain the required video footage.
We selected three videos of each target species that walked with one complete cycle (touching down to the next touching down), straight, and vertical to the camera. Each video was then converted into still images of every frame during the period between touching down and taking off using the GOM Player (GOM & Company, Seoul, South Korea). This period did not depend on time but on the target’s behavior. The convert images from the last 25% of each batch (for each measurement period) were discarded because “the muscles that are anatomically positioned to produce limb retraction—the gluteus superficialis and medius, semimembranosus and cranial biceps femoris—were active in the second half of swing and approximately the first 50–75% of stance” (Deban, Schilling & Carrier, 2012). Subsequently, the first 75% of the stance phase of each step for every specimen was divided into 12 equal time periods (particularly for each step) to obtain 13 images, including the first and last frames (Fig. 2A). The following lines were then drawn on each of the 13 pictures using Inkscape (Inkscape project) and the angle between them was measured: a line between the ankle joint and the proximal end of the tibia parallel to the Achilles tendon, and a line between the ischial tuberosity and the proximal end of the tibia (Fig. 2B).
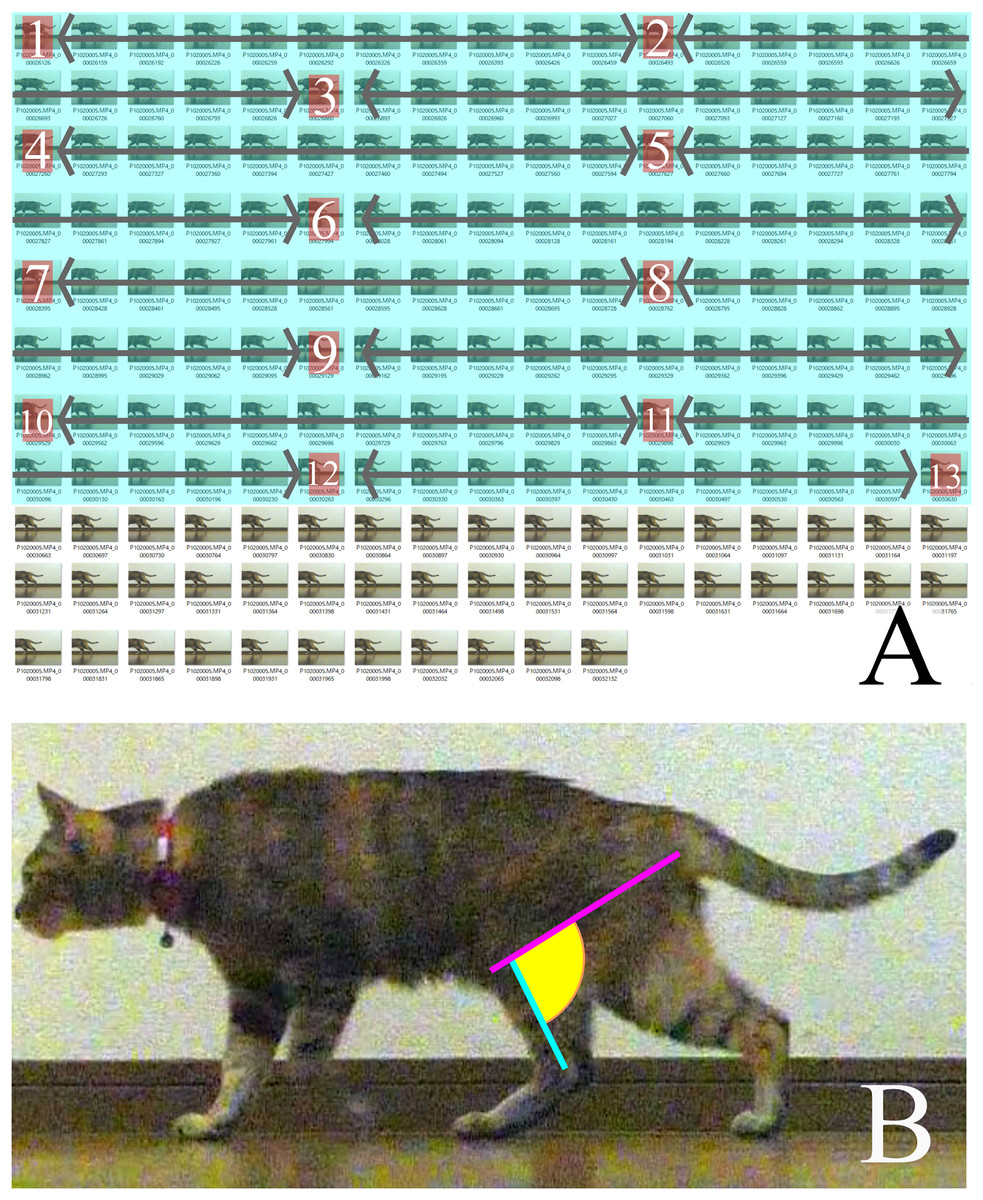
Figure 2: (A) How the measured pictures were picked.
(B) The line of measured angle on walking cat. (A) A period between touched down to the next touched down was converted to still images. The pictures covered in blue were the first 75% of the total still images. Calculated from the number of first 75% to obtain 13 pictures to measure from the first and the last combined at 12 equal intervals. The numbers in each box are measured pictures. The calculation for intervals is: (number of first 75% of still images)/(13-1) = Interval. (B) A pink line represents a line of action of m. semimembranosus. A blue line represents line of axis of tibia. A yellow area is an angle where is measured.{kind=link}
We defined each image of the 13 pictures as a “stance instance” (SI) and numbered them as SI-1 to SI-13. The combination of these 13 images defined a series of a single stance. We measured the joint angle between the lines in each of the 13 images, in each stance, and three stances for each target species in this way. The average angle of each SI was defined as θsm−t. The body mass of each species was obtained from the literature (Table 1) or zoo records. We compared the transition of θsm−t in a stance among species and locomotor modes (unguligrade, digitigrade, and plantigrade), and the average θsm−t values (i.e., θave) against body mass. Statistical analyses were performed using the R software package (R Core Team, 2020). We calculated the standard deviation (SD) to compare the variance of θsm−t among taxa, SIs, and locomotor modes. We also calculated correlation coefficient (r) to examine relationships between body mass, and θave and performed analysis of variance (ANOVA) to clarify the relationships of θave with body mass, taxa and locomotor modes. For the comparison between θave and body mass, the studied species were divided into the following groups: <1, <10, <100, <1,000, and ≥1,000 kg. The orders and locomotor modes used for the analyses were the same as in the tables. In addition, data that had only one taxon were eliminated, specifically Elephas, Dolichotis, and Macropus in the analysis between taxa; Macropus in the locomotor mode comparison; and Suricata in the body mass comparison (Table 1).
Results
Six taxa, i.e., Elephas (Proboscidea), Cervus and Rangifer (Artiodactyla), Tapirus (Perissodactyla), and Felis and Panthera (Carnivora) had differences of less than 10° between the maximum and minimum angles during a stance, which means that θsm−t changed within ±5° from the middle. Of the species, Cervus had the smallest difference during a stance, 5.80° (±2.9° from the middle-value). Twelve taxa, i.e., Chlorocebus and Macaca (Primates), Dolichotis (Rodentia), Ammotragus, Capra, and Giraffa (Artiodactyla), Canis, Chrysocyon, Suricata and Helarctos (Carnivora), and Equus and Diceros (Perissodactyla), had differences between the maximum and the minimum angles during a stance of between 10° and 20°, which means that θsm−t changed within ±10° from the middle-value. Two taxa, i.e., Ursus (Carnivora), and Ceropithecus (Primates) had differences between the maximum angle and minimum angle during a stance of less than 30°, which means that θsm−t changed within ±15° from the middle-value. While Macropus (Diprotodontia) had the largest difference between the maximum and minimum angles during a stance, (31.8°), with θsm−t changing within ±16° from the middle-value, Panthera had the lowest SD (1.73°) while Macropus had the largest one (11.5°; Fig. 3 and Table 2).
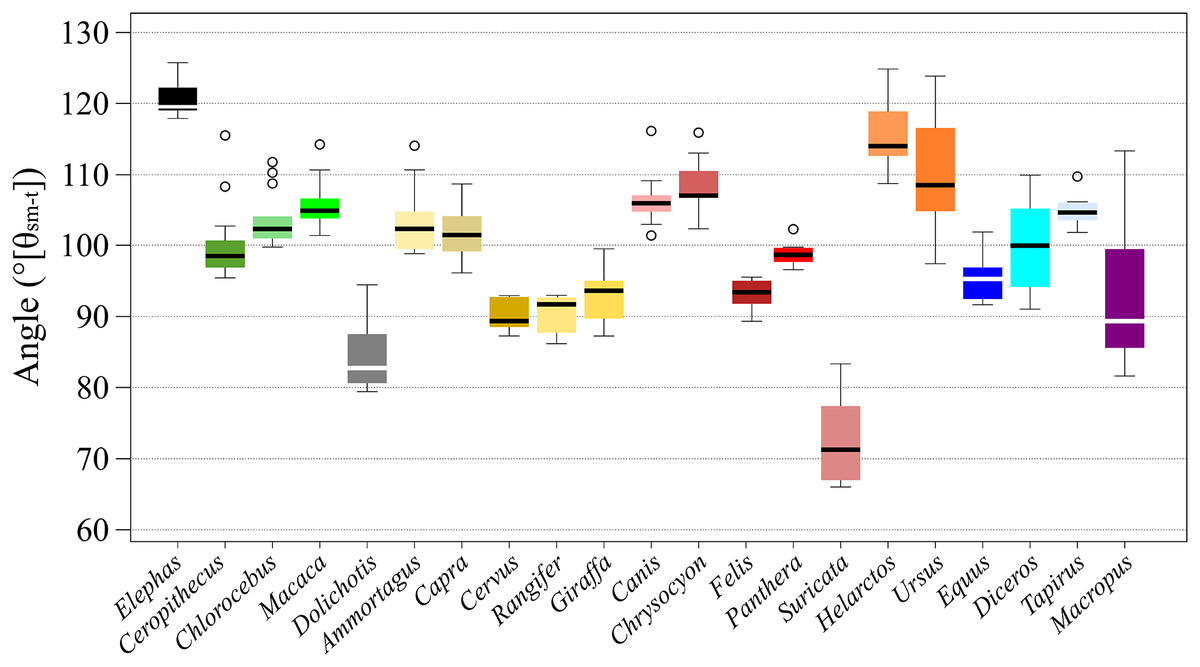
Figure 3: The box plots of θsm−t of stances of each target animals.
The x-axis shows the name of species and y-axis shows the angles in degrees. The thick bar in each box shows the median value. The top of each box shows the third quartile point, the bottom shows the first quartile point.{kind=link}
| Target | SI-1 | SI-2 | SI-3 | SI-4 | SI-5 | SI-6 | SI-7 | SI-8 | SI-9 | SI-10 | SI-11 | SI-12 | SI-13 | Diff. | θave | SD |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Proboscidea | ||||||||||||||||
| Elephas | 121.6 | 125.8 | 124.1 | 122.1 | 122.8 | 119.7 | 117.9 | 119.5 | 119.5 | 119.2 | 119.8 | 119.2 | 118.1 | 7.9 | 120.7 | 2.40 |
| Primates | ||||||||||||||||
| Cercopithecus | 115.6 | 108.4 | 102.8 | 100.2 | 100.7 | 97.7 | 98.8 | 95.6 | 95.5 | 97.0 | 96.9 | 96.3 | 98.2 | 20.1 | 100.3 | 5.64 |
| Chlorocebus | 110.3 | 102.9 | 100.6 | 100.0 | 102.4 | 104.1 | 101.7 | 101.1 | 100.4 | 102.5 | 103.5 | 108.8 | 111.9 | 11.9 | 103.9 | 3.94 |
| Macaca | 114.3 | 110.8 | 108.3 | 104.9 | 103.9 | 106.7 | 101.5 | 104.3 | 102.0 | 103.8 | 105.2 | 105.7 | 104.9 | 12.8 | 105.9 | 3.52 |
| Diff. | 5.3 | 7.9 | 7.7 | 4.9 | 3.2 | 9.0 | 2.9 | 8.7 | 6.5 | 6.8 | 8.3 | 12.5 | 13.7 | |||
| SD | 2.75 | 4.06 | 3.99 | 2.78 | 1.61 | 4.63 | 1.61 | 4.43 | 1.75 | 3.58 | 4.42 | 6.51 | 6.87 | |||
| Rodentia | ||||||||||||||||
| Dolichotis | 94.5 | 90.8 | 87.6 | 88.4 | 85.9 | 83.9 | 82.9 | 82.5 | 79.5 | 80.3 | 80.7 | 80.7 | 81.2 | 15.0 | 84.5 | 4.61 |
| Artiodactyla | ||||||||||||||||
| Ammotragus | 114.2 | 110.7 | 107.8 | 104.7 | 104.8 | 102.4 | 103.4 | 100.1 | 99.6 | 99.8 | 99.0 | 99.1 | 99.5 | 15.2 | 103.5 | 4.87 |
| Capra | 108.8 | 101.0 | 99.5 | 104.4 | 102.2 | 99.5 | 97.0 | 100.5 | 96.3 | 101.7 | 102.0 | 105.3 | 104.1 | 12.5 | 101.7 | 3.41 |
| Cervus | 93.1 | 92.8 | 92.9 | 90.7 | 89.5 | 91.1 | 89.0 | 87.9 | 87.5 | 87.3 | 88.9 | 89.3 | 93.1 | 5.8 | 90.2 | 2.18 |
| Rangifer | 93.0 | 92.8 | 93.0 | 92.7 | 92.2 | 91.7 | 91.2 | 87.9 | 89.8 | 86.3 | 87.9 | 88.1 | 92.5 | 6.7 | 90.7 | 2.39 |
| Giraffa | 99.7 | 96.7 | 95.1 | 94.9 | 94.6 | 94.9 | 93.8 | 92.6 | 90.3 | 87.3 | 89.4 | 89.8 | 88.1 | 12.4 | 92.9 | 3.65 |
| Diff. | 21.2 | 17.9 | 14.9 | 14.0 | 15.3 | 11.3 | 14.4 | 12.6 | 12.1 | 15.4 | 14.1 | 17.2 | 16.0 | |||
| SD | 9.47 | 7.44 | 6.26 | 6.63 | 6.59 | 4.92 | 5.61 | 6.23 | 5.02 | 7.57 | 6.54 | 7.55 | 6.32 | |||
| Carnivora | ||||||||||||||||
| Canis | 116.2 | 109.2 | 108.0 | 106.2 | 105.4 | 106.1 | 105.0 | 105.5 | 107.2 | 103.0 | 101.7 | 103.3 | 106.7 | 14.5 | 95.6 | 3.59 |
| Chrysocyon | 112.0 | 110.5 | 107.1 | 106.9 | 106.9 | 102.5 | 105.1 | 106.9 | 106.7 | 107.8 | 107.6 | 113.1 | 116.1 | 13.6 | 108.4 | 3.62 |
| Felis | 95.6 | 93.6 | 89.5 | 91.9 | 89.6 | 92.4 | 92.2 | 90.8 | 93.9 | 94.4 | 95.7 | 95.6 | 95.0 | 6.2 | 93.1 | 2.20 |
| Panthera | 102.4 | 99.6 | 99.1 | 99.9 | 97.9 | 97.7 | 97.8 | 98.8 | 99.2 | 98.2 | 96.6 | 98.8 | 102.5 | 5.9 | 99.1 | 1.73 |
| Suricata | 83.5 | 82.0 | 79.6 | 77.3 | 75.5 | 71.3 | 73.1 | 71.0 | 68.8 | 66.9 | 67.1 | 66.7 | 66.0 | 17.5 | 73.0 | 6.09 |
| Helarctos | 125.0 | 122.1 | 118.8 | 112.9 | 114.6 | 108.8 | 110.6 | 113.3 | 110.4 | 113.2 | 114.2 | 115.5 | 120.8 | 16.2 | 115.4 | 4.89 |
| Ursus | 124.0 | 120.8 | 117.1 | 116.5 | 113.3 | 110.1 | 108.6 | 108.1 | 105.1 | 104.9 | 101.9 | 100.4 | 97.5 | 26.5 | 109.9 | 8.08 |
| Diff. | 41.5 | 40.1 | 39.2 | 39.2 | 39.1 | 18.8 | 37.5 | 42.3 | 41.6 | 46.3 | 47.1 | 48.8 | 54.8 | |||
| SD | 15.32 | 14.59 | 14.34 | 13.46 | 14.02 | 13.48 | 13.02 | 14.38 | 14.32 | 15.18 | 14.97 | 16.08 | 17.93 | |||
| Perissodactyla | ||||||||||||||||
| Equus | 102.8 | 99.7 | 99.5 | 96.8 | 96.2 | 95.3 | 95.6 | 95.3 | 92.5 | 91.7 | 92.0 | 92.4 | 92.5 | 11.1 | 95.6 | 3.32 |
| Diceros | 110.0 | 109.9 | 106.5 | 105.3 | 103.7 | 104.8 | 98.6 | 100.0 | 95.4 | 94.3 | 91.1 | 91.1 | 91.5 | 18.9 | 100.2 | 7.01 |
| Tapirus | 109.9 | 106.0 | 103.6 | 106.0 | 106.3 | 106.3 | 104.5 | 105.1 | 104.9 | 102.2 | 102.0 | 102.0 | 104.8 | 7.9 | 104.9 | 2.19 |
| Diff. | 7.2 | 10.2 | 7.0 | 9.2 | 10.1 | 11.0 | 8.9 | 9.8 | 12.4 | 10.5 | 10.9 | 10.9 | 13.3 | |||
| SD | 4.55 | 5.13 | 3.51 | 5.09 | 5.22 | 5.96 | 4.51 | 4.90 | 6.45 | 5.43 | 6.03 | 5.92 | 7.38 | |||
| Diprotodontia | ||||||||||||||||
| Macropus | 113.4 | 113.6 | 109.0 | 99.4 | 93.2 | 85.7 | 84.1 | 81.8 | 82.1 | 87.6 | 86.4 | 92.5 | 89.4 | 31.8 | 93.7 | 11.50 |
| Total | ||||||||||||||||
| Diff. | 41.5 | 43.8 | 44.5 | 44.8 | 47.3 | 48.4 | 44.8 | 48.5 | 50.7 | 52.3 | 52.7 | 52.5 | 54.8 | |||
| Mean | 107.6 | 104.7 | 102.4 | 101.1 | 100.1 | 98.7 | 97.7 | 97.6 | 96.5 | 96.6 | 96.6 | 97.8 | 98.8 | 99.7 | ||
| SD | 11.18 | 11.18 | 10.73 | 10.03 | 10.64 | 10.61 | 10.22 | 11.33 | 11.39 | 11.66 | 11.56 | 12.05 | 12.81 |
Note:
The angle data are the average of three stances collected in our study. The differences between the maximum and the minimum angle were calculated (Diff). To visualize the variability, the standard deviation of each row and column was also calculated (SD). The raw data are in the appendix.
Based on the differences between each SI among all target species, SI-1 had the smallest difference at 41.5°, and SI-13 had the largest difference at 54.8°. However, the smallest SD was observed for SI-4 (10.03°), while the largest statistically significant SD was for SI-13, (12.81°; Table 2). This is because the low θsm−t value for Suricata is considered as an outlier in SI-13 (Fig. 4). Taxonomically, Carnivora had the greatest difference between the largest and smallest angles for the same SI, being 54.8° in SI-13; this order had relatively high differences compared to the other taxa at every SI, exceeding 30° in each case (Table 2). The smallest difference was observed in Primates, being 2.9° in SI-7; this order had relatively low differences compared to the other taxa in nine out of the 13 SIs (Table 2). Based on locomotion, digitigrade species have higher difference in SI-11 (52.7°; Table 3), while digitigrade had relatively high differences in all SIs, exceeding 38° in every case. The differences for unguligrade and plantigrade fell between 11.8° and 23.3° (Table 3). Except for Elephas and Macropus, all of the examined species had positive values when θsm−t of SI-2 was subtracted from SI-1, while when subtracting the values SI-2 from SI-3 values were positive for all species except Cervus and Rangifer. This indicates that these species, Cervus, Rangifer, Elephas and Macropus, started their stance phase by flexing the knee joint. The number of species with negative values increase in the subsequent steps, but the values soon became positive. The subtracted values of successive SIs were repeatedly positive and negative with in short span up to SI-9 and most species presented negative values after SI-10, showing extension of the knee joint when finishing the stance phase. The difference between successive SIs did not exceed 10° in any species, therefore, θsm−t smoothly transited and changed in small amounts during a stance phase (Table 4).
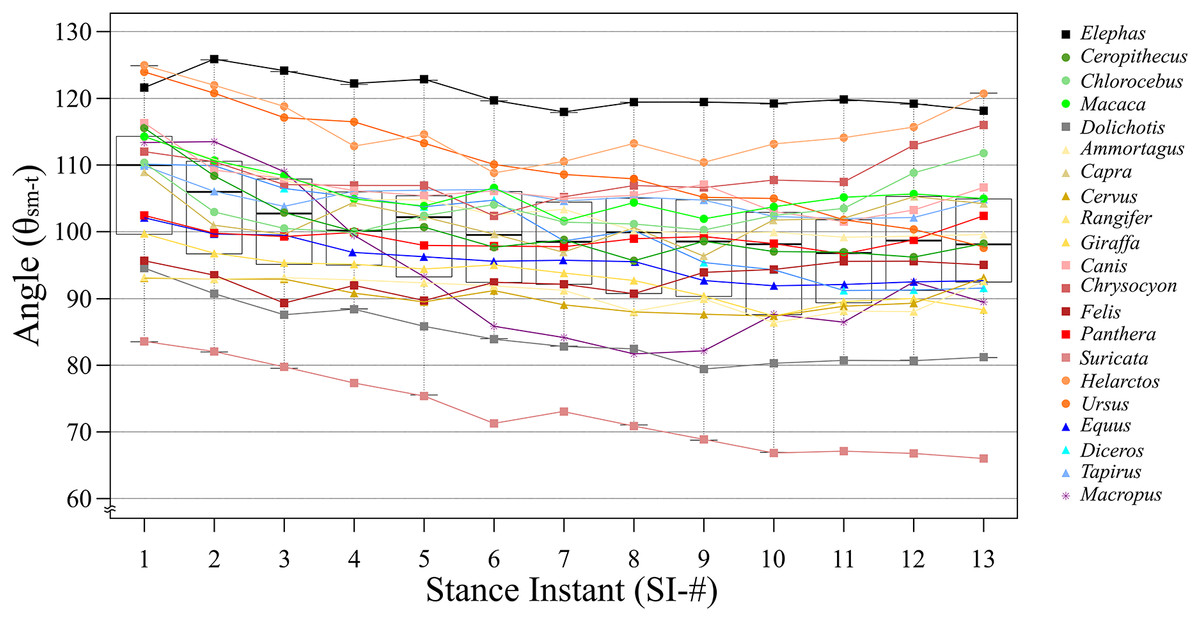
Figure 4: The line and box charts of θsm−t transition of stances of each target animals.
The x-axis shows SIs and the y-axis shows the θsm−t in degrees. Each line and plot color are identical to the colors of each box in Fig. 2. The circle plots showed the plantigrades, the triangle plots show the unguligrade, the square plots show the digitigrades, and an asterisk show other walk style. The top and bottom bars showed the maximum angles and the smallest angles, respectively. The plots under bars are outliers.{kind=link}
| Locomotor mode | SI-1 | SI-2 | SI-3 | SI-4 | SI-5 | SI-6 | SI-7 | SI-8 | SI-9 | SI-10 | SI-11 | SI-12 | SI-13 | θave | SD |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Unguligrade | |||||||||||||||
| Ammotragus | 114.2 | 110.7 | 107.8 | 104.7 | 104.8 | 102.4 | 103.4 | 100.1 | 99.6 | 99.8 | 99.0 | 99.1 | 99.5 | 103.5 | 4.87 |
| Capra | 108.8 | 101.0 | 99.5 | 104.4 | 102.2 | 99.5 | 97.0 | 100.5 | 96.3 | 101.7 | 102.0 | 105.3 | 104.1 | 101.7 | 3.41 |
| Cervus | 93.1 | 92.8 | 92.9 | 90.7 | 89.5 | 91.1 | 89.0 | 87.9 | 87.5 | 87.3 | 88.9 | 89.3 | 93.1 | 90.2 | 2.18 |
| Rangifer | 93.0 | 92.8 | 93.0 | 92.7 | 92.2 | 91.7 | 91.2 | 87.9 | 89.8 | 86.3 | 87.9 | 88.1 | 92.5 | 90.7 | 2.39 |
| Giraffa | 99.7 | 96.7 | 95.1 | 94.9 | 94.6 | 94.9 | 93.8 | 92.6 | 90.3 | 87.3 | 89.4 | 89.8 | 88.1 | 92.9 | 3.65 |
| Equus | 102.8 | 99.7 | 99.5 | 96.8 | 96.2 | 95.3 | 95.6 | 95.3 | 92.5 | 91.7 | 92.0 | 92.4 | 92.5 | 95.6 | 3.32 |
| Diceros | 110.0 | 109.9 | 106.5 | 105.3 | 103.7 | 104.8 | 98.6 | 100.0 | 95.4 | 94.3 | 91.1 | 91.1 | 91.5 | 100.2 | 7.01 |
| Tapirus | 109.9 | 106.0 | 103.6 | 106.0 | 106.3 | 106.3 | 104.5 | 105.1 | 104.9 | 102.2 | 102.0 | 102.0 | 104.8 | 104.9 | 2.19 |
| Diff. | 21.25 | 17.9 | 14.9 | 15.3 | 16.8 | 15.2 | 15.5 | 17.2 | 17.4 | 15.9 | 14.1 | 17.2 | 16.7 | ||
| SD | 8.10 | 7.08 | 5.84 | 6.31 | 6.37 | 5.85 | 5.45 | 6.31 | 5.72 | 6.68 | 5.96 | 6.55 | 6.22 | ||
| Digitigrade | |||||||||||||||
| Elephas | 121.6 | 125.8 | 124.1 | 122.1 | 122.8 | 119.7 | 117.9 | 119.5 | 119.5 | 119.2 | 119.8 | 119.2 | 118.1 | 120.7 | 2.40 |
| Dolichotis | 94.5 | 90.8 | 87.6 | 88.4 | 85.9 | 83.9 | 82.9 | 82.5 | 79.5 | 80.3 | 80.7 | 80.7 | 81.2 | 84.5 | 4.61 |
| Canis | 116.2 | 109.2 | 108.0 | 106.2 | 105.4 | 106.1 | 105.0 | 105.5 | 107.2 | 103.0 | 101.7 | 103.3 | 106.7 | 95.6 | 3.59 |
| Chrysocyon | 112.0 | 110.5 | 107.1 | 106.9 | 106.9 | 102.5 | 105.1 | 106.9 | 106.7 | 107.8 | 107.6 | 113.1 | 116.1 | 108.4 | 3.62 |
| Felis | 95.6 | 93.6 | 89.5 | 91.9 | 89.6 | 92.4 | 92.2 | 90.8 | 93.9 | 94.4 | 95.7 | 95.6 | 95.0 | 93.1 | 2.20 |
| Panthera | 102.4 | 99.6 | 99.1 | 99.9 | 97.9 | 97.7 | 97.8 | 98.8 | 99.2 | 98.2 | 96.6 | 98.8 | 102.5 | 99.1 | 1.73 |
| Suricata | 83.5 | 82.0 | 79.6 | 77.3 | 75.5 | 71.3 | 73.1 | 71.0 | 68.8 | 66.9 | 67.1 | 66.7 | 66.0 | 73.0 | 6.09 |
| Diff. | 38.1 | 43.3 | 44.5 | 44.8 | 47.3 | 48.4 | 44.8 | 48.5 | 50.7 | 52.3 | 52.7 | 52.5 | 52.1 | ||
| SD | 13.56 | 14.69 | 15.12 | 14.63 | 15.67 | 15.68 | 15.04 | 16.33 | 17.39 | 17.44 | 17.33 | 18.18 | 18.92 | ||
| Plantigrade | |||||||||||||||
| Cercopithecus | 115.6 | 108.4 | 102.8 | 100.2 | 100.7 | 97.7 | 98.8 | 95.6 | 95.5 | 97.0 | 96.9 | 96.3 | 98.2 | 100.3 | 5.64 |
| Chlorocebus | 110.3 | 102.9 | 100.6 | 100.0 | 102.4 | 104.1 | 101.7 | 101.1 | 100.4 | 102.5 | 103.5 | 108.8 | 111.9 | 103.9 | 3.94 |
| Macaca | 114.3 | 110.8 | 108.3 | 104.9 | 103.9 | 106.7 | 101.5 | 104.3 | 102.0 | 103.8 | 105.2 | 105.7 | 104.9 | 105.9 | 3.52 |
| Helarctos | 125.0 | 122.1 | 118.8 | 112.9 | 114.6 | 108.8 | 110.6 | 113.3 | 110.4 | 113.2 | 114.2 | 115.5 | 120.8 | 115.4 | 4.89 |
| Ursus | 124.0 | 120.8 | 117.1 | 116.5 | 113.3 | 110.1 | 108.6 | 108.1 | 105.1 | 104.9 | 101.9 | 100.4 | 97.5 | 109.9 | 8.08 |
| Diff. | 14.7 | 19.2 | 18.2 | 16.5 | 13.9 | 12.4 | 11.8 | 17.7 | 14.9 | 16.2 | 17.3 | 19.2 | 23.3 | ||
| SD | 8.10 | 7.08 | 5.84 | 6.31 | 6.37 | 5.85 | 5.45 | 6.31 | 5.72 | 6.68 | 5.96 | 6.55 | 6.22 | ||
| Other | |||||||||||||||
| Macropus | 113.4 | 113.6 | 109.0 | 99.4 | 93.2 | 85.7 | 84.1 | 81.8 | 82.1 | 87.6 | 86.4 | 92.5 | 89.4 | 93.7 | 11.50 |
Note:
Data for each SI angle are the same as in Table 2. The θave are the average values of each stance (a series of SI-1 to SI-13). The differences between the maximum and the minimum angle were calculated (Diff). To visualize the variability, the standard deviation of each row and column was also calculated (SD). The raw data are in the appendix.
| Target | SI1–SI2 | SI2–SI3 | SI3–SI4 | SI4–SI5 | SI5–SI6 | SI6–SI7 | SI7–SI8 | SI8–SI9 | SI9–SI10 | SI10–SI11 | SI11–SI12 | SI12–SI13 | Locomotor mode |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Elephas | −4.25 | 1.71 | 1.97 | −0.66 | 3.11 | 1.75 | −1.52 | 0.00 | 0.29 | −0.64 | 0.62 | 1.05 | D |
| Cercopithecus | 7.21 | 5.62 | 2.61 | −0.52 | 2.97 | −1.08 | 3.23 | −2.97 | 1.50 | 0.19 | 0.53 | −1.83 | P |
| Chlorocebus | 7.42 | 2.33 | 0.61 | −2.46 | −1.67 | 2.43 | 0.57 | 0.71 | −2.14 | −0.98 | −5.32 | −3.07 | P |
| Macaca | 3.49 | 2.51 | 3.44 | 0.97 | −2.81 | 5.19 | −2.82 | 2.30 | −1.73 | −1.46 | −0.48 | 0.78 | P |
| Dolichotis | 3.73 | 3.20 | −0.86 | 2.47 | 2.00 | 1.07 | 0.35 | 3.02 | −0.81 | −0.34 | −0.05 | −0.46 | D |
| Ammotragus | 3.48 | 2.88 | 3.07 | −0.10 | 2.40 | −0.94 | 3.30 | 0.51 | −0.23 | 0.81 | −0.19 | −0.32 | U |
| Capra | 7.79 | 1.53 | −4.93 | 2.26 | 2.66 | 2.51 | −3.47 | 4.19 | −5.39 | −0.28 | −3.33 | 1.20 | U |
| Cervus | 0.28 | −0.06 | 2.19 | 1.15 | −1.60 | 2.13 | 1.13 | 0.36 | 0.22 | −1.57 | −0.43 | −3.76 | U |
| Rangifer | 0.19 | −0.22 | 0.31 | 0.55 | 0.47 | 0.52 | 3.30 | −1.95 | 3.51 | −1.54 | −0.19 | −4.42 | U |
| Giraffa | 3.00 | 1.60 | 0.19 | 0.69 | −0.67 | 1.14 | 1.17 | 2.30 | 2.98 | −2.11 | −0.37 | 1.76 | U |
| Canis | 6.97 | 1.29 | 1.71 | 0.80 | −0.62 | 1.03 | −0.46 | −1.68 | 4.16 | 1.35 | −1.64 | −3.42 | D |
| Chrysocyon | 1.49 | 3.41 | 0.13 | −0.02 | 4.49 | −2.67 | −1.8 | 0.24 | −1.12 | 0.24 | −5.56 | −3.01 | D |
| Felis | 2.03 | 4.02 | −2.36 | 2.32 | −2.84 | 0.24 | 1.40 | −3.08 | −0.51 | −1.29 | 0.08 | 0.55 | D |
| Panthera | 2.79 | 0.49 | −0.75 | 2.02 | 0.16 | −0.07 | −1.00 | −0.41 | 0.96 | 1.58 | −2.12 | −3.76 | D |
| Suricata | 1.56 | 2.41 | 2.25 | 1.85 | 4.18 | −1.85 | 2.14 | 2.21 | 1.92 | −0.22 | 0.44 | 0.66 | D |
| Helarctos | 2.91 | 3.30 | 5.90 | −1.65 | 5.76 | −1.80 | −2.70 | 2.89 | −2.87 | −0.94 | −1.33 | −5.30 | P |
| Ursus | 3.19 | 3.66 | 0.58 | 3.24 | 3.22 | 1.46 | 0.56 | 2.92 | 0.23 | 3.02 | 1.50 | 2.87 | P |
| Equus | 2.36 | 0.18 | 2.73 | 0.63 | 0.88 | −0.32 | 0.29 | 2.78 | 0.82 | −0.31 | −0.41 | −0.07 | U |
| Diceros | 0.16 | 3.36 | 1.21 | 1.61 | −1.15 | 6.27 | −1.43 | 4.57 | 1.15 | 3.19 | −0.05 | −0.37 | U |
| Tapirus | 3.90 | 2.43 | −2.37 | −0.29 | 0.01 | 1.76 | −0.64 | 0.27 | 2.71 | 0.17 | −0.01 | −2.79 | U |
| Macropus | −0.20 | 4.55 | 9.63 | 6.17 | 7.47 | 1.63 | 2.27 | −0.33 | −5.38 | 1.14 | −6.10 | 3.07 | O |
Note:
Each cell shows the subtracted value of θsm−t. The italics cells show negative values. Locomotor mode abbreviations are the same as in Table 1. The bold cells show positive values.
According to the results of the θsm−t transition analysis, every studied species had relatively small differences between maximum and minimum θsm−t values during the stance phase (Figs. 3, 4, and Table 2). This showed that the total stance differences among the target animals were small; thus, θave values were representative of each species. Accordingly, we analyzed the relationships between θave and body mass. The resulting correlation coefficient (r) for all target animals was 0.30 with a p-value of 0.19 and 19 degrees of freedom (d.f.; Table 5). The correlation between the body mass and θave of each taxon was also calculated, which was significant only for Carnivora (r = 0.81, p = 0.028, d.f. = 5). The correlation between body mass and θave for each locomotor mode was only significant for digitigrade (r = 0.88, p = 0.01, d.f. = 5; Table 5). Thus, there was no statistically significant correlation between θave and body mass except for Carnivora and digitigrade. Furthermore, the θave of all species was 99.7°, with the smallest being that of Suricata (73.0°), with the largest that of Elephas (120.7°). Therefore, more than 80% of the targets (17/21) had an angle between 90° and 110° (Table 2) including all Artiodactyla, Perissodactyla, and five of the seven Carnivora assessed in our study.
| Body mass of X | Correlation coefficient | p-value (<0.05) | Degrees of freedom |
|---|---|---|---|
| All | 0.19 | 0.55 | 19 |
| Primates | 0.81 | 0.40 | 1 |
| Artiodactyla | −0.46 | 0.44 | 3 |
| Carnivora | 0.81 | 0.028 | 5 |
| Perissodactyla | 0.71 | 0.49 | 1 |
| Plantigrade | 0.82 | 0.09 | 3 |
| Digitigrade | 0.88 | 0.01 | 5 |
| Unguligrade | 0.07 | 0.87 | 6 |
ANOVAs of θave values were used to compare taxa, locomotor mode, and body mass. Only locomotor mode was statistically significant (p = 0.049; Table 6A). Furthermore, the multiple comparisons among locomotor modes showed a significant difference between unguligrade and plantigrade species (p = 0.04; Table 6B).
| A. One-way ANOVA | ||||
|---|---|---|---|---|
| Explanatory variable | F-value | Numerator d.f. | Denominator d.f. | p-value |
| Taxa | 1.62 | 3.00 | 6.82 | 0.27 |
| Ambulatory style | 4.16 | 2.00 | 9.80 | 0.049 |
| Body mass | 0.67 | 3.00 | 6.03 | 0.60 |
| B. Multiple comparisons | ||||
|---|---|---|---|---|
| Pair | Mean diff. | Standard error | d.f. | p-value |
| Unguligrade vs digitigrade | 0.44 | 6.38 | 7.40 | 0.997 |
| Unguligrade vs plantigrade | 9.65 | 3.29 | 8.77 | 0.041 |
| Digitigrade vs plantigrade | −9.21 | 6.55 | 7.97 | 0.383 |
Notes:
List (A) shows results of one-way ANOVA. List (B) shows results of multiple comparisons.
d.f., degree of freedom; diff., difference.
Discussion
Quadrupedal animals use their limbs for inverted pendulum-like movements (Cavagna, Heglund & Taylor, 1977; Griffin, Main & Farley, 2004). Physically, the swing velocity depends on the rod length; in terrestrial mammals, the swing speed affects the walking velocity. In this regard, limbs are the only structure that control the distance between the ground and body trunk; therefore, the rod length depends on the joint angles. The knee joint receives forces to flex from several influencing factors, such as the collision at touchdown, gravity, and a rising the center of mass. This means that the extensor muscles react immediately against flexion. In addition, quadrupedal mammals recovered up to 70% of their mechanical energy to lift and accelerate their center of mass via an inverted pendulum mechanism (Griffin, Main & Farley, 2004). Therefore, the joint angle should also be maintained constant to keep the length of the pendulum arm. Co-contraction also occurs to increase joint stiffness (Hogan, 1984), i.e., flexor and extensor muscles are stimulated at the same time. Previous studies focused on the position of the femur while, in our study, we center the attention on the location of the ischial tuberosity of the pelvis. As the pelvis does not rotate drastically during walking, this logic also applies to θsm−t. Each examined stance showed different results than our expectation, that extension and flexion periods were not completely separated as in the case of extension in the first half of a stance and flexion in the later half. The difference between θsm−t in successive SIs showed that joint flexion and extension were repeated over a short timespan (Table 4). The alternating increase and decrease in θsm−t between each SI allow quadrupedal mammals to maintain joint angles. In other words, the role of co-contraction during walking is not to fix the joint angles but to maintain the joint angles within a certain range, involving small increases and decreases in θsm-t across the broad range of studied taxa (Table 4). Therefore, the θsm-t angle transitions during one stance were small among the target species. Furthermore, the angle transition waveforms resembled among studied species (Fig. 4). Macropus had a unique waveform because they support their body with a tail and move both hindlimbs together (O’Connor et al., 2014). This walking pattern was found only in Macropus. Although there are different waveforms, we found that 18 out of 21 target species had only slight differences in θsm−t change (less than ±10° from the middle value) even though the largest difference was ±15.86° (Figs. 3, 4 and Table 2).
The θave values of most of the studied species (>80%) were 100 ± 10° (i.e., excluded species are Elephas, Dolichotis, Helarctos and Suricata; Table 2 and Fig. 5) including those of all three locomotor modes (i.e., unguligrade, digitigrade, and plantigrade), and five out of seven orders (i.e., Primates, Artiodactyla, Carnivora, Perissodactyla, and Marsupialia), with slight differences between unguligrade and plantigrade (p = 0.04; Table 6B and Fig. 6). Species within this range also had a wide range of the body masses, from 4.8 kg (Felis) to 1,100 kg (Diceros; Tables 1 and 3). The effective mechanical advantage (EMA) is one means of estimating mammalian limb posture; the larger EMA, the more upright the posture, with the largest species typically having greater EMA (Biewener, 1989, 1990, 2005; Dick & Clemente, 2017). Even when for the new measurement proposed in our work a slight correlation can be observed between the knee angle and body mass (Fig. 5), this correlation is not significant (r = 0.3 and p = 0.19, d.f. = 19), when considering all studied species. Also, our findings show that θave is much less variable than EMA. Such a difference between studies is due to the differences in angle-measurement positions. The ischial tuberosity, to which the m. semimembranosus is attached, is located near the posterior end of the pelvis. The horizontal or vertical orientation of the pelvis is related to body mass, with a larger body mass having a more upright orientation (Polly, 2007). Therefore, a larger body mass has a larger difference between the angle of the femur-tibia (the traditional knee joint angle, as previously standardized) than the m. semimembranosus-tibia (θsm−t and θave, as proposed in our study). In other words, θem−t and θave have the advantage of reflecting the small differences between these angles in large-body-mass species and small body mass species. Furthermore, the EMA does not increase linearly above species weighing 300 kg (Biewener, 1990, 2005; Dick & Clemente, 2017), and felids have a crouched posture even with a larger body mass (Day & Jayne, 2007; Dick & Clemente, 2017). In contrast, θave shows constant values (100 ± 10°) among all locomotor modes and a wide body mass range (4.5–1,100 kg; Tables 1 and 3).
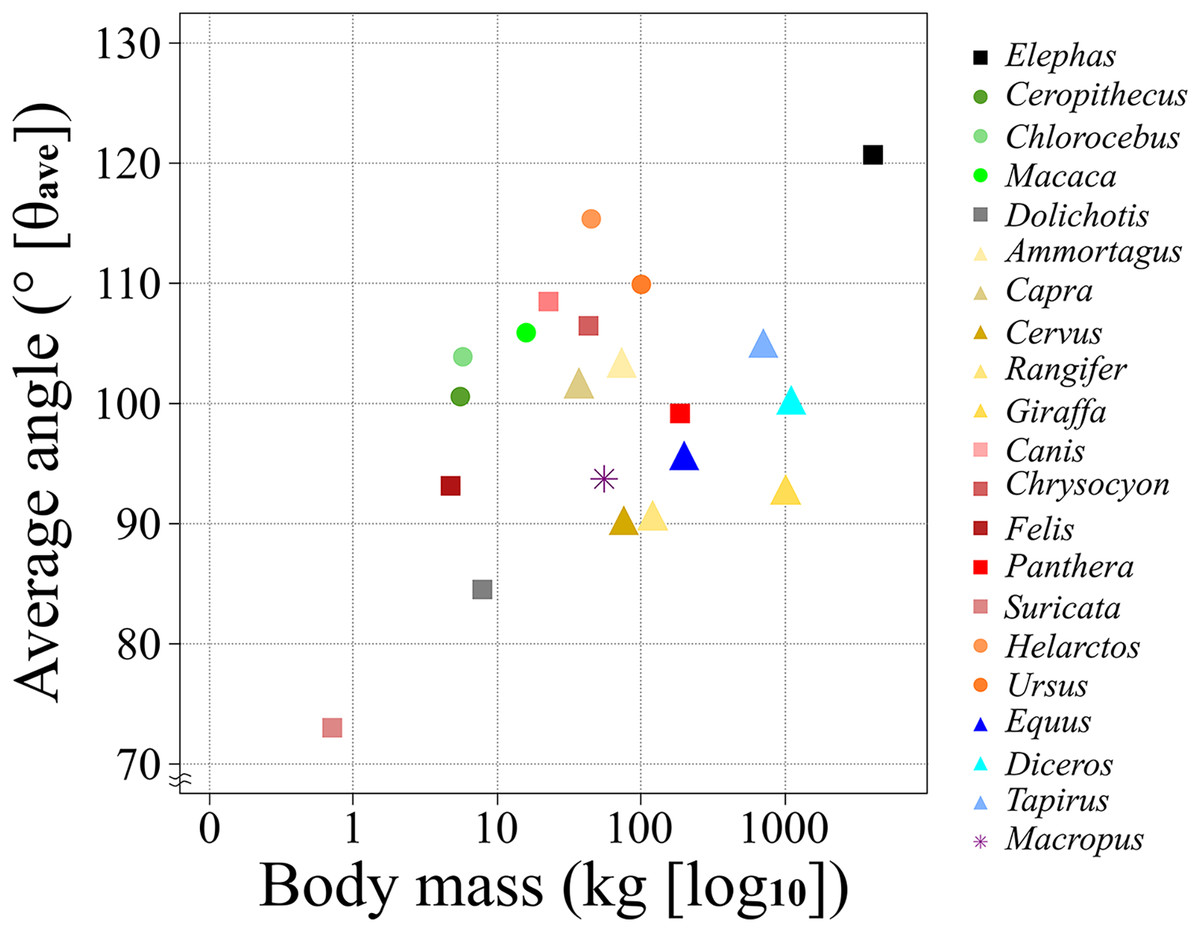
Figure 5: A scatter chart of the body mass and θave.
The x-axis shows the body mass in the log of kg and the y-axis shows the θave. Each plot color is identical to the colors of each box in Fig. 2. The circle plots showed the plantigrades, the triangle plots show the unguligrade, the square plots show the digitigrades, and an asterisk shows another walk style.{kind=link}
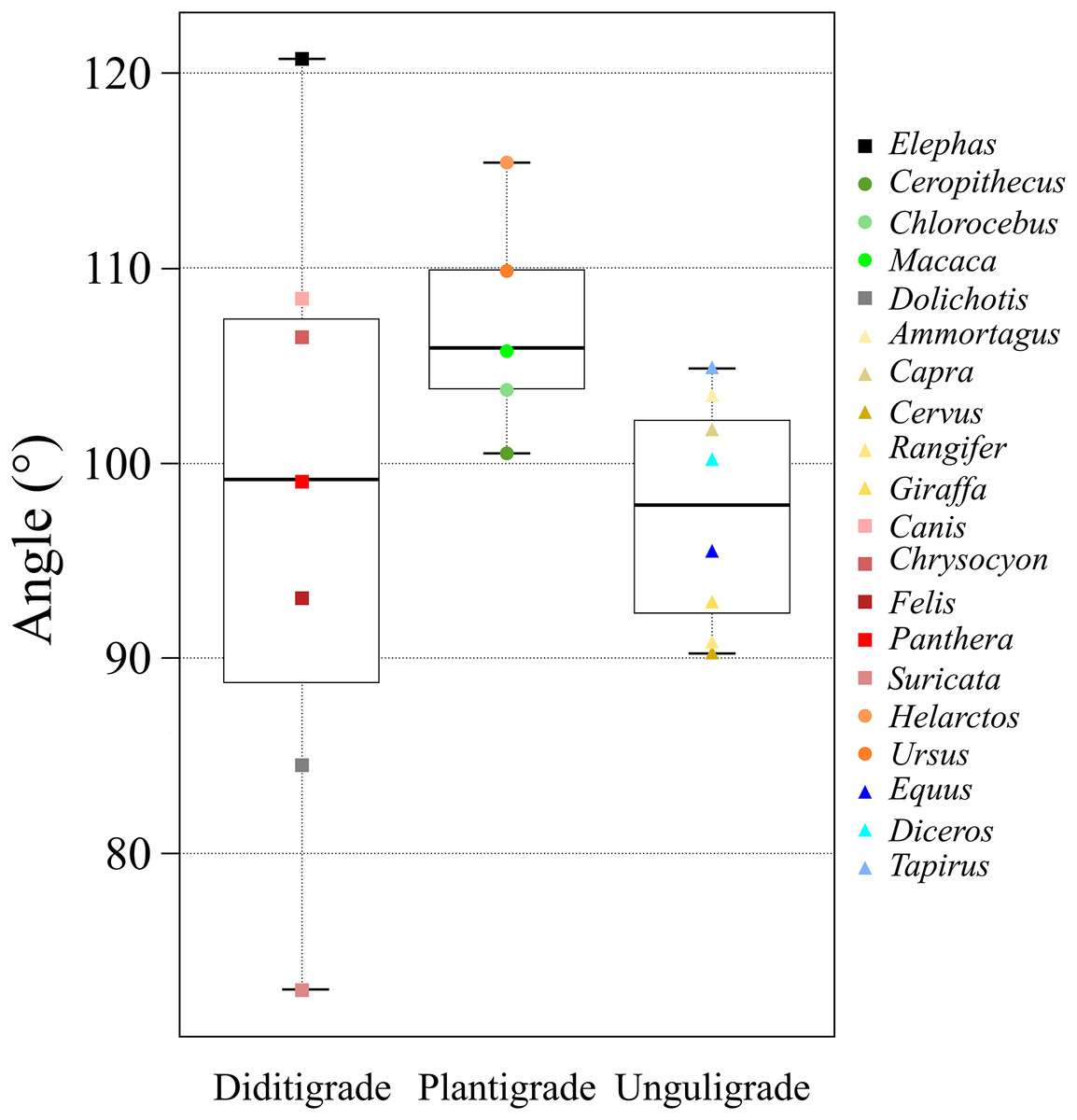
Figure 6: A box plot of θave of each ambulatory style.
The x-axis shows the locomotor modes and the y-axis shows the θave. Each plot color is identical to the colors of each box in Fig. 2. The circle plots showed the plantigrades, the triangle plots show the unguligrade, the square plots show the digitigrades.{kind=link}
In addition, we measured θsm−t based on three points on the skeleton the ischial tuberosity interior-proximal end of the tibia, and distal end of the tibia (Fig. 1). This indicates that the position of the ischial tuberosity and tibia can be fixed with 100 ± 10° on extant terrestrial quadrupedal mammals, including those with no closely related extant descendants. If a femur exists or its shape can be estimated, the limb posture can be reconstructed with higher accuracy using our approach because both the caput femoris and the distal end of the femur can be placed in the determined positions, which are the acetabulum and the proximal end of the tibia, respectively. For example, Desmostylia has been previously reconstructed in several different postures even whole-body skeletons exist (Shikama, 1966; Inuzuka, 1988; Domning, 2002; Inuzuka, Sawamura & Watabe, 2006). Because this extinct mammal has no closely related descendants and has an extremely unusual tibia, the distal half of the tibia is strongly medially twisted by approximately about 40° (Shikama, 1966; Inuzuka, 1988), no extant mammals have tibias resembling to those of Desmostylia. The θave value, which is 100 ± 10°, is not strongly affected by taxonomy, body mass, and locomotor mode, and therefore, this degree can be applied to Desmostylia.
Conclusion
Stimulation of the agonist and antagonist muscles, known as co-contraction, increases joint stiffness. In the case of the knee joint angle, our result show that θsm−t transition shown as almost flat wave-form; θsm−t did not change drastically during the first 75% of SIs during the stance phase, and the co-contraction associated with by part of the m. quadriceps femoris and the m. semimembranosus seems to effectively supports the constant posture of the knee joint in most terrestrial mammals. More than 80% of the target animals in our study had similar θave (100 ± 10°) including species across a wide range of taxa, body masses, and locomotor modes, θsm−t is measured from three points on the skeletons. Our findings indicate that θave can be a useful criteria for reconstructing the joint angles and posture of extinct mammals even if they have no closely related extant descendants.
The correlation between body mass and θave by taxon, and the angles unique to taxon and locomotor modes suggest the possibility of applying a correction to 100 ± 10° that could be applied to the all mammals. However, because our study focused on examining trends across a wide range of taxa, the sample size for each taxon was small. Further data collection and validation are required to obtain more accurate values for such corrections. In particular, our results showed a significant difference between unguligrade and plantigrade, therefore, it would apply more accurate correction if increase data. In addition, we found that Suricata had two unique features: six out of the 13 θsm−t values were outliers when compared with the other species (Fig. 4), and the difference between θave and Dolichotis, which was the second smallest species in this study, was more than 10° (Table 2 and Fig. 5). Furthermore, the difference between Suricata and the next smallest species of Carnivora, Felis, was seven-fold in terms of body mass and 20° in terms of θave. However, the difference between Felis and the largest species of Carnivora, Ursus, was greater than 20-fold in terms of body mass but less than 20° in terms of θave (Tables 1, 2 and Fig. 5). Therefore, it is possible that the Suricata data affected the r and resulting p-value for Carnivora. This is probably because Suricata spends a lot of time underground, which limits the required height to lift the trunk and limbs of the body. As such, further data from subterranean species are necessary to confirm this hypothesis.
Supplemental Information
All measurement angles of all target animals.
Every angle measured at the position shown on Fig. 1. The average from this raw data was used in this study as the angles of each animal.