
Not so sluggish: the success of the Felimare picta complex (Gastropoda, Nudibranchia) crossing Atlantic biogeographic barriers
- Published
- Accepted
- Received
- Academic Editor
- María Ángeles Esteban
- Subject Areas
- Aquaculture, Fisheries and Fish Science, Biodiversity, Biogeography, Evolutionary Studies, Marine Biology
- Keywords
- Mid-Atlantic barrier, Mitochondrial DNA, Speciation
- Copyright
- © 2016 Almada et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2016. Not so sluggish: the success of the Felimare picta complex (Gastropoda, Nudibranchia) crossing Atlantic biogeographic barriers. PeerJ 4:e1561 https://doi.org/10.7717/peerj.1561
Abstract
The molecular phylogeny of the Atlanto-Mediterranean species of the genus Felimare, particularly those attributed to the species F. picta, was inferred using two mitochondrial markers (16S and COI). A recent revision of the Chromodorididae clarified the taxonomic relationships at the family level redefining the genus Felimare. However, conflicting taxonomic classifications have been proposed for a restrict group of taxa with overlapping morphological characteristics and geographical distributions designated here as the Felimare picta complex. Three major groups were identified: one Mediterranean and amphi-Atlantic group; a western Atlantic group and a tropical eastern Atlantic group. F. picta forms a paraphyletic group since some subspecies are more closely related with taxa traditionaly classified as independent species (e.g. F. zebra) than with other subspecies with allopatric distributions (e.g. F. picta picta and F. picta tema). Usually, nudibranchs have adhesive demersal eggs, short planktonic larval phases and low mobility as adults unless rafting on floating materials occurs. Surprisingly however, the phylogeny of the F. picta complex suggests that they successfully cross main Atlantic biogeographic barriers including the mid-Atlantic barrier. This ability to cross different biogeographic barriers may be related to F. picta’s distinct life history and ecological traits. Compared to other Chromodorididae F. picta has larger eggs and planktotrophic larvae which could be related to a longer planktonic phase.
Introduction
Nudibranchs are heterobranch mollusks (Gastropoda) that comprise more than 3000 species (Willan & Coleman, 1984). The family Chromodorididae includes some of the most striking colored nudibranchs present in almost all marine habitats. Conspicuous aposematic colorations advertise defensive adaptations that include the production or incorporation of bioactive chemicals from cnidarians or sponges upon which they feed (Wollscheid & Wägele, 1999; Haber et al., 2010). These toxins have captured the interest of numerous authors (Gaspar, Rodrigues & Calado, 2009; Haber et al., 2010; Cruz, Gaspar & Calado, 2012) and represent a source of natural compounds whose biological activity is being actively prospected. As a consequence, chromodorids are also good models for the study of color pattern evolution and mimicry in marine species (Gosliner & Johnson, 1999). Clarifying the evolutionary relationships among chromodorids would pave the way to predict characteristics that are still undescribed and would help to disentangle genetic from environmental effects (e.g. Gaspar, Rodrigues & Calado, 2009).
The taxonomy of the chromodorid nudibranchs was originally based on morphological data (Rudman, 1984). Gosliner & Johnson (1999) revised the phylogeny of the worldwide genus Hypselodoris, including some of the species that are more abundant in the northeastern Atlantic and Mediterranean Sea. These revisions were followed by a number of morphological (Alejandrino & Valdés, 2006) and molecular studies (Wollscheid & Wägele, 1999; Wollscheid-Lengeling et al., 2001; Turner & Wilson, 2007; Johnson, 2011), including the description of several new species (Dacosta, Padula & Schrödl, 2010; Ortigosa & Valdés, 2012). Johnson & Gosliner (2012), based on two mitochondrial genes, proposed a new phylogenetic hypothesis of the chromodorid and reorganized its traditional taxonomy. These authors identified the Pacific Ocean as an effective biogeographic barrier, proposing the inclusion of all eastern Pacific, Atlantic and Mediterranean Hypselodoris, together with two species from the genus Mexichromis from the eastern Pacific and Caribbean, into a different genus, Felimare, while retaining the west/central Pacific Hypselodoris species within this genus. Eastern Pacific, Atlantic and Mediterranean species previously in the genera Chromodoris and Glossodoris were included within the Felimida. Prior to their revision, Hypselodoris, Chromodoris and Glossodoris were the most species rich chromodorid genera. Recently, using additional species of these genera and a nuclear DNA marker, Ortigosa et al. (2014) recovered a polytomy between several Atlantic and Mediterranean Felimida and Felimare species.
Taxonomic considerations of Felimare picta
Within the Chromodorididae F. picta presents the wider distribution area throughout the tropical and subtropical Atlantic Ocean, including the Mediterranean Sea and the eastern Atlantic archipelagos of Cape Verde, Canaries, Madeira and Azores (Ortea, Valdés & García-Gómez, 1996). These coastal sea slugs are one of the largest chromodorid species and vary greatly in colour and pattern, resulting in the traditional classification under many species names (e.g. Felimare (Hypselodoris) webbi (d’Orbigny, 1839) and Felimare (Hypselodoris) tema (Edmunds, 1981)). Variation in external morphology or internal anatomy also led to incongruencies in the taxonomic classification of F. picta subspecies (Ortea, Valdés & García-Gómez, 1996; Alejandrino & Valdés, 2006). One of the cases that illustrates the ambiguous situation of this group is the changing taxonomic status of the southwest Atlantic representatives classified as subspecies (Felimare picta lajensis) by Troncoso, Garcia & Urgorri (1998), as a valid species (Felimare lajensis) by Domínguez, García & Troncoso (2006) and then downgraded again to a subspecific level by Dacosta, Padula & Schrödl (2010). Similarly, Felimare tema, originally described from Ghana by Edmunds (1981) was reclassified as subspecies (Felimare picta tema) by Ortea, Valdés & Garcia-Gomez (1996). Color pattern similarities between Felimare picta verdensis and F. picta tema (sensu Ortea, Valdés & Garcia-Gomez, 1996) also raised some doubts on the validity of these subspecies. Table S1 in Supplementary Data summarizes the main taxonomic classifications and also the overlapping distribution areas of some of these subspecies.
The phylogenetic relationships of the F. picta complex with other taxa long recognised as distinct species, both from the east and west Atlantic, remains largely unknown. F. picta is the only amphi-Atlantic species of this genus, but along the east Atlantic shores and central Atlantic islands we can find several species with overlapping distributions. Along the west Atlantic shore this phylogenetic relationship is even more complex, with a close trans-isthmian phylogenetic relationship between all eastern Pacific and the Atlantic/Mediterranean Felimare species, together with two species previously assigned to the genus Mexichromis (F. porterae and F. kempfi) reported by Johnson & Gosliner (2012).
Biogeography
The ability to cross biogeographic barriers is normally restricted to highly mobile species or species that produce propagules with high potential to disperse at least during a particular phase of their life-cycle (Briggs, 1974). Aside from landmasses, long extensions of deep oceanic water and abrupt changes of physical or chemical properties of marine water can effectively restrict the colonization potential of inshore organisms (Floeter et al., 2008; Luiz et al., 2011). Furthermore, the effectiveness of these permeable barriers may be influenced by the potential to establish new populations in the recently colonized habitats (Luiz et al., 2011).
The mid Atlantic barrier (MAB) is a deep ocean gap formed by the Atlantic Ocean basin in the last 85 Myr. It spans a long distance with a minimum straight-line distance of approximately 2800 km, therefore acting as a barrier to larval dispersal for marine organisms (Floeter et al., 2008; Luiz et al., 2011).
For molluscs, Vermeij (2005) concluded that 30.8% (northeastern Atlantic) to 47.3% (northwestern Atlantic) of the North-Atlantic species present an amphi-Atlantic distribution. However, F. picta is the Atlanto-Mediterranean Chromodorididae with the wider distribution area (Ortea, Valdés & García-Gómez, 1996) and one of the few present in both margins of the Atlantic. the Mediterranean and most islands of the tropical and temperate north Atlantic, having crossed major biogeographic barriers. With such an extensive geographical distribution, several subspecies were historically assigned to F. picta. Some of these subspecies are morphologically very similar and present overlapping geographical distributions (e.g. F. picta picta and F. picta webbi cf. Ortea, Valdés & García-Gómez, 1996). A molecular phylogeny of this group would shed some light on the taxonomic status of F. picta subspecies and other closely related Atlanto-Mediterranean Felimare species.
In the present study, we intend to clarify the taxonomic status of F. picta and its relationships with other congeneric species. Simultaneously, we want to evaluate the validity of its previously proposed subspecies that currently raise numerous identification problems to many taxonomists. The molecular phylogeny will be the starting point to infer the biogeographic relationships within a group of taxa that was able to cross the main Atlantic biogeographic barriers.
Material and Methods
Sampling
The species sampled in the present study, the geographical origin of the samples and the GenBank accession numbers are listed in Table S2 in the Supplementary Data. Specimens were identified and portions of tissue were provided by a nudibranch taxonomist (Dr. Gonçalo Calado–see Gaspar, Rodrigues & Calado (2009); Coelho & Calado (2010); Haber et al. (2010); Calado & Silva (2012); Cruz, Gaspar & Calado (2012)). The same nudibranch taxonomist kindly provided pictures of several Felimare species and F. picta subspecies (Supplementary Data). Extracted DNA is available from ISPA laboratory collections. Voucher specimens of F. lajensis are available from Museu de Zoologia da Universidade de São Paulo (MZSP97468). In an attempt to detect possible intraspecific variability in this species, a total of 32 F. picta representing samples from its entire geographical distribution area and either side of the MAB were analysed. Additional sampling included the Strait of Gibraltar and the Mauritanian cold water barrier limiting dispersal in other marine species (e.g. the Strait of Gibraltar) (see Patarnello, Volckaert & Castilho, 2007). Samples belonging to 13 different Felimare species were also collected. These represent approximately half of the described species of the eastern Pacific, Atlantic and Mediterranean (WoRMS Editorial Board, 2014), including some already available in GenBank, which were used for comparative purposes to provide a broader phylogenetic framework and access the overall genetic divergence within this genus.
DNA extraction, amplification and sequencing
DNA was extracted from tissue samples preserved in ethanol, using a proteinase K/SDS based protocol (Sambrook, Fritsch & Maniatis, 1989). Primers used to amplify a fragment 461 bp long from the 16S mitochondrial rDNA (16Ssar 5′ CGC CTG TTT ATC AAA AAC AT 3′ and 16Sbr 5′ CCG GTC TGA ACT CAG ATC ACG T 3′), and a fragment 554 bp long from the COI (LCO1490 5′ GGT CAA CAA ATC ATA AAG ATA TTG G 3′ and HCO2198 5′ TAA ACT TCA GGG TGA CCA AAA AAT CA 3′), are described in Palumbi et al. (1996) & Folmer et al. (1994), respectively. The primers 28SC1(F) 5′ ACC CGC TGA ATT TAA GCA T 3′ and 28SD3(R) 5′ GAC GAT CGA TTT GCA CGT CA 3′ used by Mollaret et al. (1997), Vonnemann et al. (2005) & Klussmann-Kolb et al. (2008) were also used to attempt to amplify a fragment of the nuclear 28S rDNA in these species.
PCR conditions were conducted as follows: 2 min. 95 °C followed by 35 cycles of (95 °C (30 sec.), 54 °C (30 sec.) and 72 °C (60 sec.)) for the 16S fragment; 2 min. 95 °C followed by 35 cycles of (95 °C (45 sec.), 50 °C (45 sec.) and 72 °C (60 sec.)) for the COI fragment and 4 min. 95 °C followed by 38 cycles of (94 °C (30 sec.), 52 °C (50 sec.) and 72 °C (120 sec.)) followed by 10 min. at 72 °C for the 28S fragment.
PCR products were purified using microClean (MicroZone, www.microzone.co.uk), and sequenced in STABVIDA (http://www.stabvida.net/) using these primers.
Phylogenetic analysis
Following Johnson (2011) we used an outgroup species from the Dendrodorididae family: Doriopsilla pelseneeri d’Oliveira, 1895. DNA sequences were analysed and edited using the software program CodonCode aligner (v. 3.5, CodonCode Corporation) and were aligned separately using M-Coffee (Notredame, Higgins & Heringa, 2000). Manual alignment masking was performed, by excluding loci scored as ‘bad’ by M-Coffee, in order to improve sign-to-noise ratio. Transitional saturation was examined by plotting transitions and transversions against sequence divergence using GTR distance and implementing Xia et al. test (Xia et al., 2003; Xia & Lemey, 2009) test of substitution saturation available in Dambe v. 5.3.108 (Xia, 2013).
Each fragment and a concatenation of both fragments were analysed using four phylogenetic inference methods: 1) maximum-parsimony (MP) with 100 heuristic searches using random additions of sequences and implementing the TBR algorithm, as implemented in PAUP 4.0b10 (Swofford, 2001); 2) minimum-evolution (ME), also implemented in PAUP with 1000 resamplings, was implemented using the best-fit model of molecular evolution chosen according the Bayesian Information Criterion as implemented in JModeltest 2.0 (Darriba et al., 2012); 3) Maximum Likelihood, as implemented in RaxML (Stamatakis, Hoover & Rougemont, 2008) and 4) Bayesian inference (BI) performed using MCMC as implemented in MrBayes v. 3.2 (Ronquist et al., 2012), with two independent runs of four Metropolis-coupled chains of four million generations each in order to estimate the posterior probability distribution. Topologies were sampled every 100 generations and a majority-rule consensus tree was estimated after discarding the first 10% samples. Convergence was verified by inspecting the average standard deviation of split frequencies and tracing likelihood throughout samples in Tracer v1.6 (Drummond et al., 2012). Both ML and BI analyses of the concatenated alignment considered two partitions for which independent parameters were estimated. For the first three phylogenetic inference methods, branch support values for each node were tested by bootstrap analysis, with 100 resamplings (Felsenstein, 1985). Net between group mean distances were calculated using Mega (Tamura et al., 2013) using Tamura-Nei distance with gamma model estimated by the composite likelihood method.
Results
Sequence analysis
The null hypothesis of congruence between the two data sets (16S and COI rDNA) was not rejected (P = 0.33) by the ILD test (Farris et al., 1995). Therefore, the results presented in subsequent sections relate to the analysis of the concatenation of the 16S and COI rDNA fragments. The combined sequence of the 16S + COI rDNA fragments resulted in an alignment of 1026 base pairs. Of these, 712 characters are constant, 66 variable characters are parsimony-uninformative and 248 are parsimony-informative characters. No saturation was observed for the 16S and COI datasets or the concatenated fragment with both sequences (P < 0.001 for all combinations) (Xia et al., 2003; Xia & Lemey, 2009).
Minimum-evolution (ME), using the best-fit model of molecular evolution chosen according the Bayesian Information Criterion as implemented in JModeltest 2.0 (Darriba et al., 2012) was HKY + I + G for 16S; TrN + I + G for COI and TIM3 + I + G for the concatenated 16S and COI.
Since we were not able to amplify the COI fragment of F. acriba, known from the Caribbean, this species is not shown in the concatenated tree with the 16S and COI fragments (Fig. 1). However, all phylogenetic analysis with the 16S fragment recovered this species as the sister species of F. bayeri with very high support values (Bayesian analysis with posterior probability of 1.0 and maximum parsimony with bootstrap value of 100).
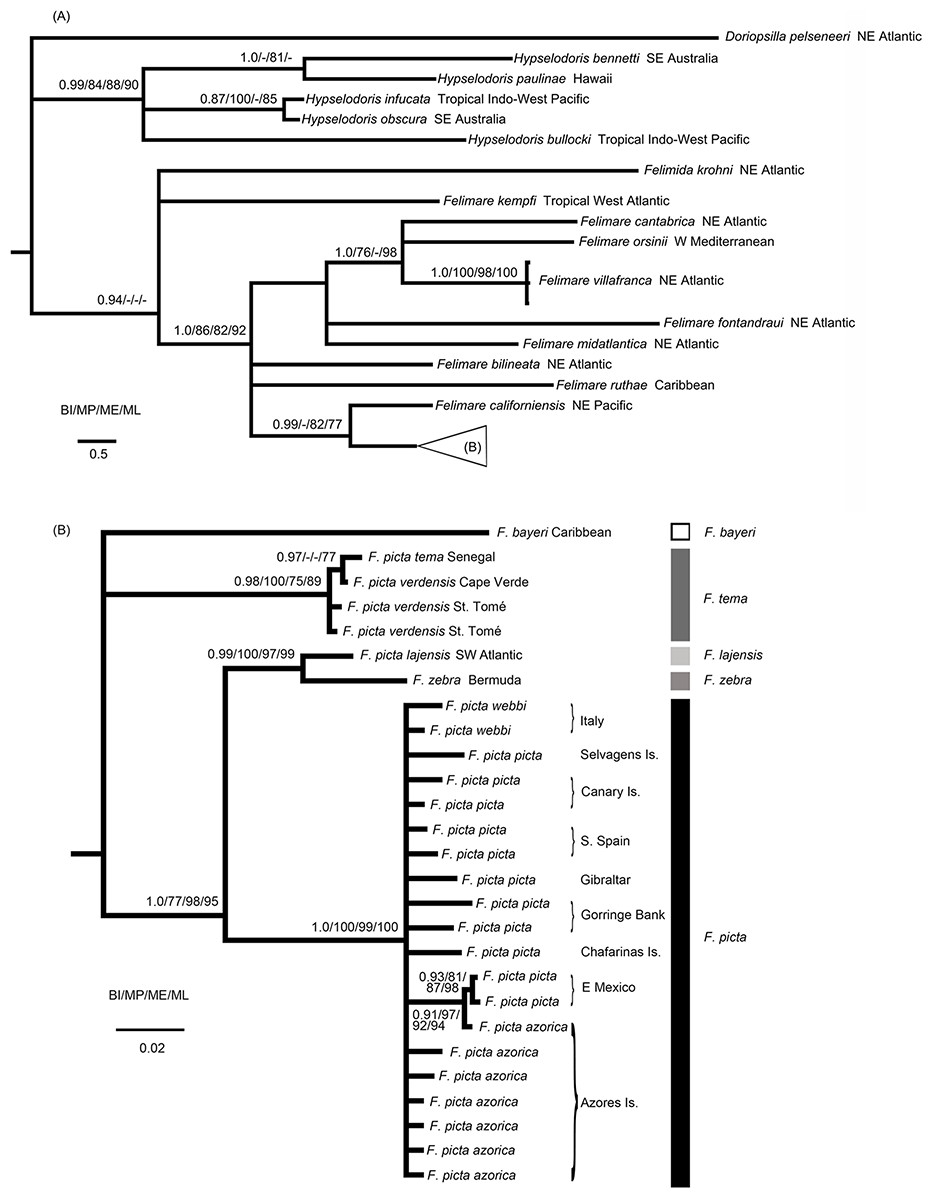
Figure 1: Phylogenetic relationships of the Atlanto-Mediterranean “F. picta complex” and other related species.
(A) Phylogenetic relationships between several Indo-West Pacific Hypselodoris species and eastern Pacific, Atlantic and Mediterranean Felimare species obtained from the 16S + COI molecular data. Bayesian posterior probabilities (BI) and MP, ME and ML bootstrap support are shown above and below each node, respectively. (B) Phylogenetic relationships of the Atlanto-Mediterranean “F. picta complex.”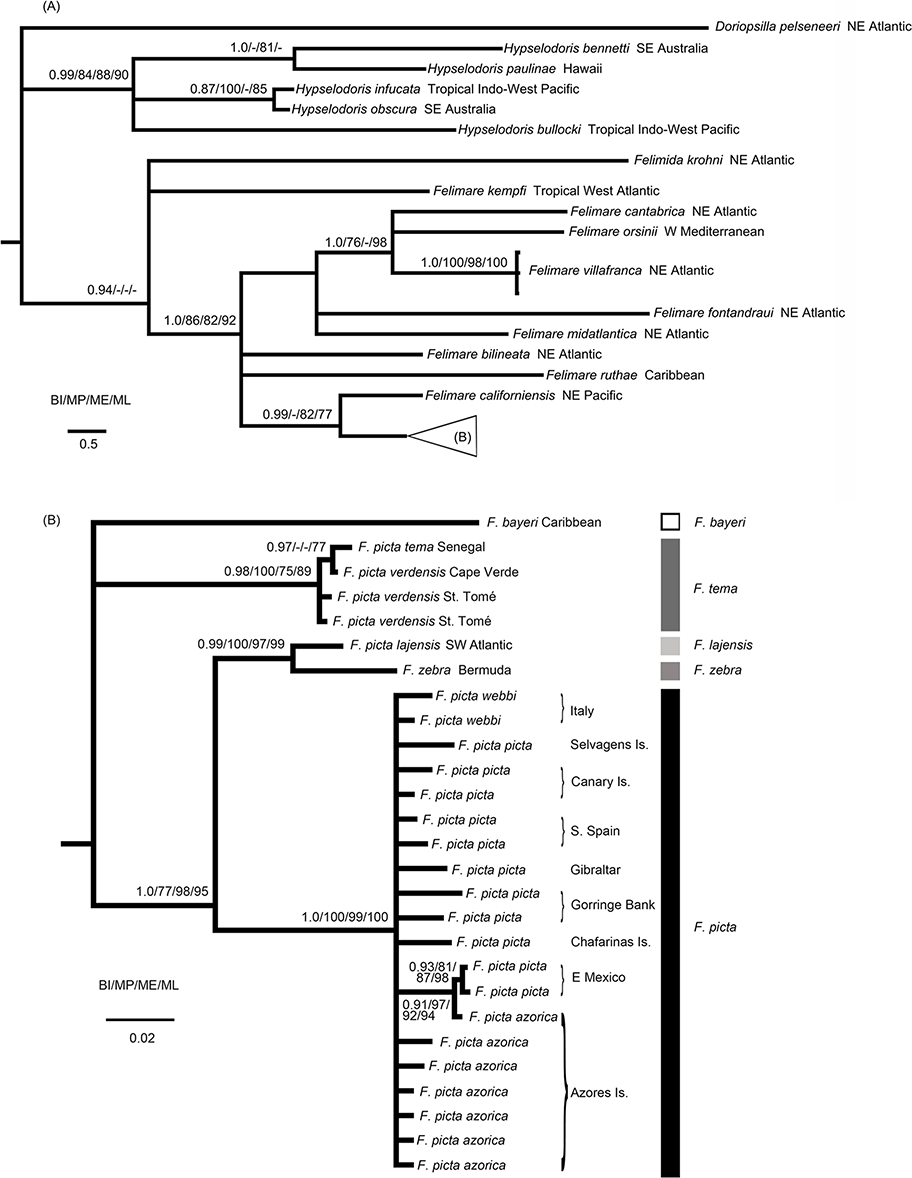{kind=link}
Estimated net evolutionary divergence between species and subspecies is presented in Table 1. Genetic similarities and haplotypes shared between F. picta collected from Mexico to the Mediterranean, including the Azores (F. picta picta, F. picta webbi and F. picta azorica sensu Ortea, Valdés & García-Gómez, 1996) revealed no genetic isolation. The genetic distance between these subspecies and the west African samples (0.255–0.272), which would be classified as F. p. tema and F. p. verdensis (sensu Ortea, Valdés & García-Gómez, 1996) is larger than the distance shown for F. zebra (0.185) and is similar to the one shown for F. bayeri (0.277).
| Chromodoris | F. bayeri | F. bilineata | F. californensis | F. cantabrica | F. fontadraui | F. kempfi | F. lajensis | F. midatlantica | F. orsinii | F. picta azorica | F. picta picta | F. picta webbi | F. picta tema | F. picta verdensis | F. ruthae | F. villafranca | F. zebra | Hypselodoris | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chromodoris | 0,546 | 0,617 | 0,613 | 0,735 | 0,631 | 0,496 | 0,628 | 0,560 | 0,624 | 0,690 | 0,702 | 0,691 | 0,565 | 0,560 | 0,872 | 0,576 | 0,591 | 0,625 | |
| F. bayeri | 1,766 | 0,284 | 0,218 | 0,470 | 0,395 | 0,489 | 0,079 | 0,307 | 0,389 | 0,082 | 0,084 | 0,083 | 0,067 | 0,063 | 0,582 | 0,370 | 0,082 | 0,614 | |
| F. bilineata | 2,055 | 0,922 | 0,250 | 0,464 | 0,379 | 0,561 | 0,288 | 0,286 | 0,371 | 0,289 | 0,290 | 0,273 | 0,278 | 0,269 | 0,502 | 0,420 | 0,300 | 0,623 | |
| F. californensis | 2,069 | 0,692 | 0,771 | 0,442 | 0,363 | 0,648 | 0,199 | 0,330 | 0,382 | 0,197 | 0,197 | 0,192 | 0,182 | 0,179 | 0,630 | 0,363 | 0,209 | 0,769 | |
| F. cantabrica | 2,592 | 1,536 | 1,520 | 1,454 | 0,478 | 0,786 | 0,449 | 0,461 | 0,405 | 0,422 | 0,432 | 0,427 | 0,444 | 0,429 | 0,747 | 0,280 | 0,494 | 0,910 | |
| F. fontadraui | 2,095 | 1,260 | 1,227 | 1,162 | 1,596 | 0,633 | 0,453 | 0,309 | 0,329 | 0,481 | 0,479 | 0,480 | 0,395 | 0,381 | 0,602 | 0,399 | 0,441 | 0,700 | |
| F. kempfi | 1,660 | 1,611 | 1,865 | 2,211 | 2,752 | 2,217 | 0,519 | 0,586 | 0,653 | 0,576 | 0,577 | 0,572 | 0,533 | 0,522 | 0,845 | 0,679 | 0,542 | 0,702 | |
| F. lajensis | 2,138 | 0,258 | 0,930 | 0,644 | 1,485 | 1,499 | 1,749 | 0,338 | 0,458 | 0,050 | 0,050 | 0,050 | 0,074 | 0,074 | 0,652 | 0,346 | 0,022 | 0,749 | |
| F. midatlantica | 1,839 | 0,941 | 0,919 | 1,053 | 1,497 | 0,968 | 1,973 | 1,085 | 0,356 | 0,362 | 0,365 | 0,370 | 0,316 | 0,320 | 0,614 | 0,270 | 0,373 | 0,674 | |
| F. orsinii | 2,044 | 1,194 | 1,175 | 1,206 | 1,373 | 1,059 | 2,208 | 1,489 | 1,147 | 0,487 | 0,490 | 0,484 | 0,381 | 0,386 | 0,627 | 0,206 | 0,440 | 0,689 | |
| F. picta azorica | 2,340 | 0,272 | 0,944 | 0,627 | 1,364 | 1,572 | 1,966 | 0,163 | 1,130 | 1,579 | 0,000 | 0,000 | 0,081 | 0,078 | 0,651 | 0,408 | 0,054 | 0,734 | |
| F. picta picta | 2,379 | 0,277 | 0,949 | 0,628 | 1,405 | 1,564 | 1,973 | 0,165 | 1,138 | 1,591 | 0,000 | 0,000 | 0,083 | 0,080 | 0,647 | 0,412 | 0,053 | 0,732 | |
| F. picta webbi | 2,338 | 0,272 | 0,891 | 0,617 | 1,387 | 1,569 | 1,957 | 0,164 | 1,155 | 1,569 | 0,000 | 0,000 | 0,084 | 0,080 | 0,636 | 0,404 | 0,055 | 0,709 | |
| F. picta tema | 1,847 | 0,211 | 0,885 | 0,572 | 1,450 | 1,259 | 1,798 | 0,246 | 0,994 | 1,180 | 0,262 | 0,269 | 0,272 | 0,004 | 0,614 | 0,404 | 0,089 | 0,690 | |
| F. picta verdensis | 1,826 | 0,200 | 0,866 | 0,570 | 1,406 | 1,207 | 1,767 | 0,250 | 1,019 | 1,218 | 0,255 | 0,259 | 0,260 | 0,008 | 0,600 | 0,397 | 0,090 | 0,687 | |
| F. ruthae | 3,040 | 1,936 | 1,691 | 2,161 | 2,538 | 2,027 | 2,945 | 2,218 | 2,051 | 2,156 | 2,212 | 2,206 | 2,161 | 2,090 | 2,035 | 0,620 | 0,614 | 0,850 | |
| F. villafranca | 1,920 | 1,128 | 1,313 | 1,124 | 0,901 | 1,274 | 2,339 | 1,068 | 0,830 | 0,635 | 1,269 | 1,283 | 1,256 | 1,263 | 1,247 | 2,112 | 0,362 | 0,636 | |
| F. zebra | 1,942 | 0,258 | 0,943 | 0,650 | 1,617 | 1,441 | 1,841 | 0,072 | 1,192 | 1,388 | 0,181 | 0,179 | 0,185 | 0,281 | 0,287 | 2,045 | 1,100 | 0,701 | |
| Hypselodoris | 2,209 | 2,135 | 2,113 | 2,736 | 3,266 | 2,459 | 2,473 | 2,693 | 2,350 | 2,401 | 2,585 | 2,577 | 2,484 | 2,423 | 2,424 | 2,978 | 2,182 | 2,463 |
Phylogenetic analysis
The genus Felimare and higher order relationships
The results presented in Fig. 1A support the distinctiveness of the eastern Pacific, Atlantic and Mediterranean Felimare species compared to the Indo-Pacific Hypselodoris species. The Caribbean Felimare kempfi failed to be included in a distinct clade with the remaining species of this genus.
The Felimare picta complex
The monophyly of F. picta, including specimens from all geographical areas, is not supported given the internal position of several west Atlantic species, such as F. zebra and F. lajensis (Fig. 1B). Although ME and BI recovered a third west Atlantic taxa, F. bayeri, as the sister species of the African subspecies of F. picta (F. p. tema and F. p. verdensis), this phylogenetic relationship was not supported by ML or MP methods. As a precautionary measure these incongruent results were interpreted as an unresolved trichotomy (Fig. 1B). Nevertheless, F. bayeri was always recovered in a clade, including the remaining taxa of the F. picta complex, with very high support values. These results alone show that F. picta, as currently defined, constitutes a paraphyletic group.
The results presented in Fig. 1B also confirm that, unlike other species of the same genus, the distribution range of F. picta encompasses both margins of the Atlantic Ocean together with several North Atlantic islands, namely the Azores. Interestingly, samples from this central Atlantic archipelago had identical or similar haplotypes compared to others from Mexico (west Atlantic), Portugal and Spain (east Atlantic) or Italy (Mediterranean).
The paraphyly of F. picta and, consequently, the phylogenetic relationships between its subspecies and other related taxa requires a revision of the taxonomic status of this complex of species and raises the question of how many transatlantic colonisation events could have occurred within the F. picta complex alone.
For identification purposes, F. picta complex diagnostic nucleotide characteres are listed for 16S and COI fragments in Table S3 in the Supplementary Data.
Discussion
Molecular phylogenetic relationships suggest that current taxonomy of the genus Felimare and, in particular, the F. picta subspecies should be revised.
Currently, F. picta forms a paraphyletic group since F. lajensis, formerly classsified as F. picta lajensis, is the sister species of F. zebra, another west Atlantic Felimare species. West African F. tema, formerly described as an F. picta subspecies, should be reinstated as an independent species. F. picta, the only chomodorid with a distribution area encompassing both margins of the Atlantic Ocean, including the Archipelago of the Azores, and the Mediterranean Sea, includes former F. picta picta, F picta webbi and F. picta azorica. No genetic structure was detected between these putative subspecies. The taxonomic and biogeographic implications of these proposals are addressed in the sections below.
Taxonomic implications
Although presenting a distinct color pattern when compared to other F. picta, the genetic data presented here suggests that the Azorean samples are not isolated from the west or east Atlantic and Mediterraneam samples. Although differences in phenotype may result from recent genetic divergence or distinct ecological influences, the classification of taxa into different species or subspecies should be accompanied by genetic studies that are able to demonstrate their monophyly. The results are compatible with two alternative hypotheses: (i) genetic isolation between F. picta subspecies was recent, requiring fast evolving genetic markers, such as microsatellites, to detect reproductive isolation or; (ii) at least some of these “subspecies” coexist and interbreed. Considering the paraphyly of the F. picta complex and the fact that (Ortea, Valdés & García-Gómez, 1996) did not describe valid morphological characters or designate type specimens to justify the nomination of several new subspecies (e.g. F. p. azorica), F. p. picta, F. p. webbi or F. p. azorica should, by order of precedence and for the time being, be synonymized under F. picta (Schultz, 1836) instead of assigning them to different subspecies. This is particularly important within the chromodorids, which are known to be quite diversified in terms of color patterns. In fact, some of the characters described for some subspecies, such as the absence of yellow lines in F. p. azorica, are also observed in F. p. picta and the lack of discriminating differences is even more evident between the latter and F. p. webbi.
F. p. tema (Edmunds, 1981) and F. p. verdensis (Ortea, Valdés & García-Gómez, 1996) show no genetic differences between samples collected from Senegal and from Cape Verde. These results are not surprising since (Ortea, Valdés & Garcia-Gómez, 1996) reclassified F. tema, originally described by Edmunds (1981) from Ghana as a subspecies of F. picta, and described a new subspecies: F. p. verdensis, endemic to Cape Verde Islands, São Tomé and southern Angola. However, they did not designate any type specimens and the morphological variability described by Edmunds (1981) for F. tema is similar to that described for the specimens of F. p. tema and F. p. verdensis except for one difference related to radula secondary denticles which are absent in F. p. tema. These specimens are characterized by longitudinal orange lines, orange submarginal border, orange lined gills and a dark blue background color with lighter blue patches along the submarginal mantle border.
The results presented here suggest that, for the time being, the original taxonomic classification by Edmunds (1981) should be reinstated and F. tema should include both F. p. tema and F. p. verdensis. In fact, this tropical eastern Atlantic species, whose distribution area would now include Senegal, Ghana, south Angola and the archipelagoes of Cape Verde and São Tomé, is more distantly related to other parapatric norheastern Atlantic F. picta than the allopatric western Atlantic F. lajensis.
F. p. lajensis Troncoso, Garcia & Urgorri, (1998) was originally described for Brazil as a subspecies of F. picta but has been re-assessed as a different species by Domínguez, García & Troncoso (2006). Its ambiguous taxonomic status (species or subspecies) is rooted on the morphological descriptions that: i) distinguish this taxon from remaining F. picta, based on the dark blue to violet body pattern with blue to violet gills, five spaced dorsal lines and a deferent duct with a narrow preprostatic portion (Troncoso, Garcia & Urgorri, 1998; Domínguez, Garcia & Troncoso, 2006) or ii) suggest that it should be included in F. picta because some specimens have gill rachis with yellow lines, up to nine dorsal yellow lines and a yellow or white mantle margin (DaCosta, Padula & Schrödl, 2010). In fact, F. picta present gill rachis with yellow lines, only three dorsal lines and the preprostatic portion of the deferent duct is absent. DaCosta, Padula & Schrödl (2010) also showed that radular and allosperm receptacles differences described by Troncoso, Garcia & Urgorri (1998) & Domínguez, García & Troncoso (2006) were not supported by the additional material analysed by those authors. Due to the wide morphological variation DaCosta, Padula & Schrödl (2010) decided to keep this taxon as a subspecies of F. picta until detailed anatomical comparisons and also molecular approaches are concluded to better understand not only the satus of this taxon but also the relationships between all F. picta subspecies.
In our study, F. picta samples from Brazil were always recovered in a clade with F. zebra (Bermuda) and the genetic distance suggests a close relationship between these taxa. This southwestern Atlantic taxon is the sister clade to all remaining F. picta, excluding the east tropical Atlantic F. tema which diverged near the base of the F. picta complex as did also F. bayeri.
Although our results using two mitochondrial markers are clear and unambiguous, phylogenies based on mitochondrial DNA may result in misleading speciation histories when there are discrepancies with nuclear DNA (e.g. Zhang & Hewitt, 1996; Shaw, 2002). Joint analysis of mitochondrial and nuclear DNA may be informative even when results from both types of fragments are contradictory, particularly “when the context for the conflict is understood” (Rubinoff & Holland, 2005). In fact, nuclear phylogenies frequently reinforce mitochondrial phylogenies (e.g. Levy et al., 2011). On the other hand, nuclear loci present several technical limitations in phylogenetic inference such as low copy number, heterozygosity, paralogous loci with multiple copies and low substitution rate, which make them uninformative when comparing close related species or even subspecies (for a revision see Rubinoff & Holland, 2005). One example of this argument are the chromodorid species analysed by Ortigosa et al. (2014) where nuclear markers did not add any additional information or resolve ambiguous results on closely related Felimida species. Although, reclassifying F. picta, F. tema and F. lajensis as independent species should be considered provisional until more specimens are throughly analysed and nuclear markers are compared, the suggestions presented above clarify the taxonomy and help to define groups that are monophyletic and well characterized with mitochondrial DNA markers and morphological characters.
In a wider phylogenetic scope the results indicate a basal politomy with four branches. Western Atlantic F. ruthae and eastern Atlantic/west Mediterranean F. bilineata appear as individualized branches. A third clade includes the F. picta complex, with the amphi-Atlantic F. picta, western Atlantic F. lajensis, F. zebra and F. bayeri and eastern Atlantic F. tema and its sister species, the eastern Pacific F. californiensis. Future phylogenetic studies will show if other eastern Pacific species are included in the same or in different clades together with the remaining Atlantic and Mediterranean species. A forth clade includes the remaining eastern Atlantic and Mediterranean species: F. cantabrica, F. fontandraui, F. orsinii, F. midatlantica and F. villafranca. The exclusion of F. kempfi from the Felimare clade was already argued by Johnson & Gosliner (2012) placing this Caribbean species with the eastern Pacific F. porterae as a potential sister group to a larger clade of eastern Pacific, Atlantic and Mediterranean Felimare species (Johnson & Gosliner, 2012).
The fact that the Indo-Pacific species are included in a distinct monophyletic clade corroborates the conclusions of Johnson & Gosliner (2012) pointing to an ancient diversification of the Felimare species in the eastern Pacific, Atlantic and Mediterranean since the closure of the Tethys Sea in the East by the end of the Miocene.
Felimare picta complex biogeography
Sea surface currents and larvae characteristics, including larval behaviour, are crucial for an organism to be able to cross important biogeographic barriers. Briggs (1995) described four biogeographic provinces later reviewed by Spalding et al. (2007) & Briggs & Bowen (2012) that are relevant to this study: northeastern Atlantic Lusitania (including Macaronesia and the Mediterranean) (but see Almada et al., 2013), tropical West Africa, the Caribbean and Brazil. These provinces are delimited by several soft (non-terrestrial) barriers such as: (1) the mid-Atlantic barrier; (2) the Mauritanian cold water upwelling in the northwestern coast of Africa; (3) the Orinoco/Amazonas freshwater plume; (4) the Almeria/Oran front separating the Atlantic from the Mediterranean. Some organisms can cross these areas when environmental conditions are favourable which result in the establishment of new populations or species (e.g. Floeter et al., 2008).
Based on the phylogenetic results presented in this study the success of the F. picta complex crossing all these biogeographic barriers underlined by the amphi-Atlantic distribution of F. picta is not reflected among the other chomodorids.
Panama isthmus
The high diversity in the eastern Pacific and the opportunity to colonize the western Atlantic via Panama Isthmus before its closing, contributed to a higher species richness in this area compared to other Atlantic regions. This fact allowed not only for species to exist on both sides of the isthmus, but also some taxa to radiate to many others (Lessios, 2008). Examples of sister relationships across this particular vicariant event include opisthobranchs (e.g. Malaquias & Reid, 2009) and fish (Briggs, 1995; Muss et al., 2001; Grant, Lecomte & Bowen, 2010). This pattern is also observed between the eastern Pacific F. californiensis and the F. picta complex. In fact, the F. picta complex is more closely related with F. californiensis than with other Atlanto-Mediterranean Felimare species which probably means that the origin of this species complex is posterior to the closure of this hard (terrestrial) biogeographic barrier.
Mid-Atlantic barrier
The Atlantic ocean barrier is an important constraint to the migration of individuals between both sides of the Atlantic (Briggs, 1995). However, crossing of the MAB by some species has been suggested by the amphiatlantic distribution of these species or because they have sister taxa on both margins of the Atlantic reflecting historical speciation events (Carmona et al., 2011).
The genus Felimare and, in particular, the F. picta complex, illustrates both these patterns. Present geographic distribution of F. picta includes the west and eastern Atlantic Ocean and also the Mediterranean Sea which is supported by molecular data presented here. Although this hypothesis should be tested in the future with additional samples and appropriate phylogeographic analysis, the phylogeny of this group suggests that the mid-Atlantic barrier was probably crossed twice by this complex of species. First, an earlier colonization event resulted in the divergence of F. tema (including F. picta tema and F. picta verdensis) in equatorial Africa region. Second, a separation between the clade F. lajensis/F. zebra and F. picta (including F. picta from western and northeastern Atlantic) and a transatlantic migration resulting in the colonization of both sides of the Atlantic by this later species.
Major oceanic surface currents suggest that this migration could have followed a westward route, particularly in the equatorial region (e.g. Silva, Horne & Castilho, 2014 and references therein). With predominant surface currents from northwest Africa to Central America (North equatorial current) and from southwest Africa to southeast South America (South equatorial current), one hypothesis to explain current species distribution would be an westward migration with speciation along the American coast. However this westward migration hypothesis is highly improbable if we consider the phylogeny of this group of species, particularly the basal position of the west Atlantic species within the F. picta complex phylogeny and the eastern Pacific F. californiensis being the sister species of this clade.
An alternative hypothesis would be an eastward colonization from the western Atlantic to the European and African coasts which, if we assume that the present surface current pattern was already in effect, could follow two alternative routes: a northern route along the Gulf stream followed by the Azores and the Canaries currents (Barton, 2001) and an equatorial route following the north equatorial countercurrent (see Fonseca et al., 2004). Phylogenetic patterns showed in this work and those reported by other authors based on morphological and meristic data (Gosliner & Johnson, 1999; Alejandrino & Valdés, 2006) also support this eastward migration hypothesis.
Assuming that the origin of F. picta is posterior to the closure of the Isthmus of Panama and therefore posterior to the settlement of the Gulf Stream, the similarity between Mexican samples of F. picta and an individual collected in the Azores suggest that this archipelago may have acted, and still acts, as a stepping-stone in this northern route. This hypothesis is further supported by the fact that during glacial periods these currents were even stronger than in present times (Wunsch, 2003) which could result in the rapid transportation from west to the east Atlantic of planktotrophic larvae or adults and eggs on rafting materials. The equatorial counter current is seasonal, being stronger during the Spring (Richardson et al., 1992), when it reaches surface transport velocities between 23 cm s−1 (Richardson et al., 1992) and 45 cm s−1 (Urbano, Almeida & Nobre, 2008). This would mean that the mid Atlantic barrier could be crossed at maximum velocity in less than 10 days (38.88 km day−1). This would allow an independent eastward migration by a southern route which could have led to the origin of F. tema in the tropical west African coast. Only future phylogeographic studies with population samples of F. picta from both sides of the Atlantic may shed some light on the dispersion route of this species.
Mauritanian cold water barrier
The present allopatric distribution of F. tema and F. picta could be explained by the persistence of biogeographic barriers and/or by ecological constraints. The cold water barrier along the Mauritanian shores due to strong upwelling (Marañón et al., 2001) and the Pleistocenic glaciations could have prevented F. tema from colonizing the northeastern Atlantic shores. More recently, with the settling of F. picta in the northeastern Atlantic shores other ecological constraints may have been in effect. This argument is based on the fact that F. tema is deeply rooted within a clade with several west Atlantic extant species and F. picta shares a common ancestor with F. lajensis and F. zebra, which are also west Atlantic species. If the hypothesis of the “eastward migration” proves to be correct, present allopatric distribution of F. tema and F. picta may be the result of two independent dispersion processes: the first resulting in the speciation of the tropical east Atlantic F. tema from an west Atlantic ancestor, and the second resulting in the colonization of the northeastern Atlantic and the Mediterranean by F. picta.
Examples of this eastward migration are common across a large array of taxonomic groups and are much more common than migrations on the opposite directions (Ávila, 2005; Rocha et al., 2008; Beldade et al., 2009). This may be the result of the predominant current patterns described above and the higher species richness in the west Atlantic shores.
Orinoco/Amazonas barrier
Genetic isolation between central western and southwestern Atlantic may have occurred about 6 million years with the origin of the Amazon River (Nunan, 1992; Hoorn, 1994). Variation in salinity levels due to river fresh water plumes and the absence of appropriate hard substrate along the southwest Atlantic coast became known as the Central American Gap and represent an important biogeographic barrier for marine organisms (Rocha, 2003; Ludt & Rocha, 2014). Since the closure of the Isthmus of Panamá the patterns of epipelagic circulation remained approximately constant (Haug & Tiedemann, 1998) therefore, conditions were appropriate for this biogeographic semi-permeable barrier to promote a recent split between F. lajensis and F. zebra.
The question still remains however on why among all chromodorids has the F. picta complex experienced such a success crossing all main Atlantic biogeographic barriers? Even considering F. picta sensu strictu, this is the only chromodorid whose distribution encompasses northwestern Atlantic, northeastern Atlantic and the Mediterranean. Futhermore, nudibranchs and the chromodorids, in particular, are sedentary and have a dispersion ability that is much reduced during all life stages. Rafting on floating materials could explain their ability to colonize distant locations; however, the individuals of these species are benthic and are usually found over sponges upon which they feed. Therefore, it is improbable that these organisms may disperse during their adult phase. Coelho & Calado (2010) reported that F. picta shows the largest egg size and planktotrophic larvae length at hatching reported among nudibranch molluscs (for a review see Todd, Lambert & Davies, 2001). Although F. villafranca have even larger eggs, it presents direct and not planktotrophic development. It is commonly accepted that large larvae have higher survival rates during transport in the water column. Although, Shulman & Bermingham (1995) found no relationship between oceanographic patterns, larval duration and population genetic structuring in the Caribbean, they refer that a different scenario could emerge on larger geographic scales. For this purpose, additional studies on different species which were also able to cross the MAB, such as the Chromodorididae Tyrinna evelinae and Cadlina rumia, from a sister group of the Chromodorididae (Johnson, 2011), could be of extreme interest as well as tagging mark and recapture of these species to determine if the distance they travel enables physical crossing.
From a molecular ecology perspective, genetic assignment tests could indicate if the most likely sources of recruits are from local or distant populations (see Piry et al., 2004; Wilson & Rannala, 2003).
A comprehensive study of the phylogeny of the genus Felimare, including all its species and nuclear DNA markers is still needed to clarify the taxonomy of this group. Furthermore, phylogeographic data would provide information on the direction and number of colonization events of each taxonomic entity and would allow the implementation of species delimitation analysis (see Puillandre et al., 2012).
Nevertheless the biogeographic considerations and phylogenetic relationships described above may help to refine current information on a group of marine organisms that have been raising the attention of a broad community from evolutionary biology and ecology to natural products chemistry.
Supplemental Information
Felimare (Hypselodoris) picta subspecies.
Table S1–Distribution areas of the six subspecies described for Felimare (Hypselodoris) picta.
Sampling localities and accession numbers.
Table S2–List of material included in this study, sampling localities and Genbank accession numbers.
Diagnostic nucleotide characteres for the 16S and COI DNA fragments.
Diagnostic nucleotide characteres for the 16S and COI DNA fragments listed for F. picta (including former F. picta picta, F picta webbi and F. picta azorica), F. lajensis, F. zebra and F. tema.
Felimare picta (Mexico).
Felimare picta (Mexico). Credits to G. Calado.
{kind=link}
Felimare picta (Archipelago of the Azores).
Felimare picta (Archipelago of the Azores). Credits to G. Calado.
{kind=link}
Felimare picta (South Portugal).
Felimare picta (South Portugal). Credits to R. Coelho/IPM.
{kind=link}
Felimare tema (S. Tome and Princepe).
Felimare tema (S. Tome and Princepe). Credits to G. Calado.
{kind=link}
Felimare cantabrica (South Portugal).
Felimare cantabrica (South Portugal). Credits to R. Coelho/IPM.
{kind=link}
Felimare fontandraui (West Portugal).
Felimare fontandraui (West Portugal). Credits to G. Calado.
{kind=link}
Felimare pinna (Cape Verde).
Felimare pinna (Cape Verde). Credits to G. Calado.
{kind=link}
Felimare midatlantica (South Portugal).
Felimare midatlantica (South Portugal). Credits to R. Coelho/IPM.
{kind=link}
Felimare villafranca (South Portugal).
Felimare villafranca (South Portugal). Credits to R. Coelho/IPM.
{kind=link}
Felimare bilineata (South Portugal).
Felimare bilineata (South Portugal). Credits to R. Coelho/IPM.
{kind=link}