
New titanosauriform (Dinosauria: Sauropoda) specimens from the Upper Cretaceous Daijiaping Formation of southern China
- Published
- Accepted
- Received
- Academic Editor
- Andrew Farke
- Subject Areas
- Evolutionary Studies, Paleontology
- Keywords
- Sauropoda, Titanosauriform, Cretaceous, China
- Copyright
- © 2019 Han et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits using, remixing, and building upon the work non-commercially, as long as it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2019. New titanosauriform (Dinosauria: Sauropoda) specimens from the Upper Cretaceous Daijiaping Formation of southern China. PeerJ 7:e8237 https://doi.org/10.7717/peerj.8237
Abstract
Titanosauriform sauropod dinosaurs were once considered rare in the Upper Cretaceous of Asia, but a number of titanosauriforms from this stratigraphic interval have been discovered in China in recent years. In fact, all adequately known Cretaceous Asian sauropods are titanosauriforms, but only a few have been well studied, lending significance to any new anatomical information that can be extracted from Asia’s Cretaceous sauropod record. Here we give a detailed description of some titanosauriform bones recovered recently from the Upper Cretaceous Daijiaping Formation of Tianyuan County, Zhuzhou City, Hunan Province, southern China. The occurrence of this material in Hunan increases the known geographic range of titanosauriforms in eastern Asia. Although all of the specimens discussed in this paper can be assigned to Titanosauriformes at least tentatively, some bones display a limited number of features that are more typical of basal sauropods and/or derived diplodocoids, suggesting complex patterns of character evolution within Neosauropoda.
Introduction
By the dawn of the Cretaceous, the diversity of sauropod dinosaurs had been reduced to the major clades Diplodocoidea and Titanosauriformes, which differed greatly in their subsequent evolutionary fortunes (Wilson, 2005; Whitlock, D’Emic & Wilson, 2011; D’Emic, 2012; Fanti et al., 2015). Rebbachisaurid diplocoids underwent a modest radiation in the Early Cretaceous, and a few persisted into the early Late Cretaceous in Africa, Europe and South America, but diplodocoids were less diverse even in the Early Cretaceous than they had been in the Late Jurassic. Titanosauriforms, by contrast, achieved a worldwide distribution and a high level of diversity during the Early Cretaceous, and were among the most successful herbivorous dinosaur groups of the Late Cretaceous. Although titanosauriforms were once thought to have been relatively rare in China, it was eventually realized that all known Chinese Cretaceous sauropods were titanosauriforms (e.g., Whitlock, D’Emic & Wilson, 2011), and a series of discoveries have shown that the diversity of this group in China was substantial. To date, nine titanosauriform genera have been described from the Upper Cretaceous of China (Fig. 1), including Sonidosaurus from the Erlian Formation of Inner Mongolia (Xu et al., 2006), Borealosaurus from the Sunjiawan Formation of Liaoning Province (You et al., 2004), Huabeisaurus from the Huiquanpu Formation of Shanxi Province (Pang & Cheng, 2000), Zhuchengtitan from the Wangshi Group of Shandong Province (Mo et al., 2017), Gannansaurus from the Nanxiong Formation of Jiangxi Province (Lü et al., 2013b), Qingxiusaurus from Upper Cretaceous red beds in Guangxi Province (Mo et al., 2008), Jiangshanosaurus and Dongyangosaurus from the Jinhua Formation of Zhejiang Province (Tang et al., 2001; Lü et al., 2008; Yu et al., 2010), and Baotianmansaurus from the Xiaguan Formation of Henan Province (Zhang et al., 2009; Wang et al., 2013). The situation in China conforms to a wider biogeographic pattern, in that Titanosauriformes is the only sauropod clade known to have been present in the Cretaceous of Asia as a whole (Wilson, 2005; Wilson & Upchurch, 2009). In the Jurassic, by contrast, the sauropod fauna of China was overwhelmingly dominated by basal (non-neosauropod) taxa, the only known Chinese diplodocoid being the recently described Lingwulong from the Toarcian-Bajocian of Ningxia (Xu et al., 2018).
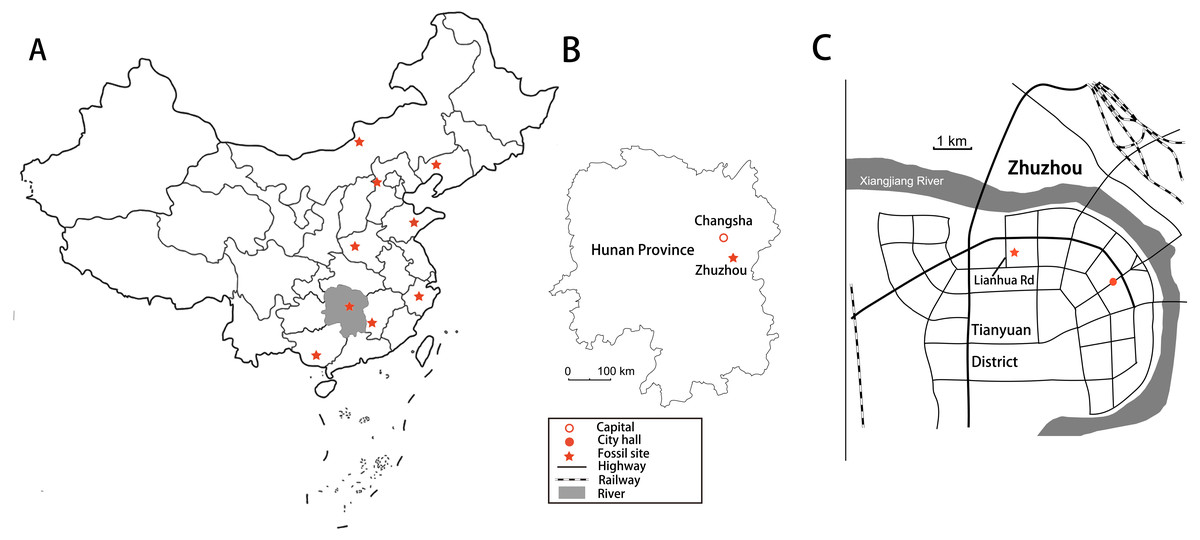
Figure 1: Locality maps showing sites that have produced sauropod dinosaurs.
(A) Map of China showing titanosauriform dinosaur localities, with Hunan Province shaded. (B) Map of Hunan Province showing the location of Zhuzhou City. (C) Position of dinosaur locality adjacent to Lianhua Road, Tianyuan District, Zhuzhou City.{kind=link}
In 2008, a new fossil site containing numerous dinosaur bones was discovered in the Upper Cretaceous Daijiaping Formation of Tianyuan County, Zhuzhou City, Hunan Province, southern China (Fig. 1). The site is on the east side of Lianhua Road, in the northern part of Tianyuan County, and was the intended location of a new middle school. Prior to construction of the school, workers were using explosives to level an area of elevated terrain when a crew member named Dangsheng Fang noticed fragments resembling animal bones among the shattered rock. Some of these fossils were sent to experts from the Institute of Vertebrate Paleontology and Paleoanthropology in Beijing, who identified them as dinosaur bones. The local government protected the site immediately and arranged for all visible fossils to be collected. More than 200 bones were gathered and apportioned between the Zhuzhou Museum and the Bureau of Land and Resources of Zhuzhou City. Most of the bones were fragmented and displaced by the explosives, and are consequently difficult to identify (Han et al., 2017) or to assign to particular beds, but some remained in place so that their stratigraphic positions could be documented as they were collected (Fig. 2).

Figure 2: Stratigraphic section through the dinosaur-bearing Daijiaping Formation of Tianyuan District, Zhuzhou City.
Modified from Han et al. (2017). Abbreviations: K2, Late Cretaceous; Q, Quaternary.{kind=link}
The dinosaur bones occurred within an area of about 6,000 m2 and within strata of the Daijiaping Formation, which includes a lower portion consisting of sandstone and an upper portion consisting of siltstone and mudstone. On a regional scale, the total thickness of the formation is about 1 km. However, the beds exposed at the site span only ∼50 m of thickness and are believed to represent a transitional region between the lower and upper parts of the formation. The exposed beds can be divided into three parts: a lower part consisting of red calcareous siltstone, interspersed with conglomerate layers; a middle part consisting of alternating siltstone and conglomerate beds, and containing the dinosaur bones; and an upper part mainly consisting of siltstone (Fig. 2). The dinosaur bones were recovered from six units in the middle part of the section, within a stratigraphic interval ranging in thickness from 8.6 to 16.0 m across the width of the outcrop (Fig. 2). The Daijiaping Formation is generally considered to date from the early Late Cretaceous (Coniacian to Santonian) (Han et al., 2017). However, analysis of a total of 22 pollen samples from layers 4 and 9, situated in the middle part of the exposed section, instead suggests a latest Cretaceous (Campanian to Maastrichtian) age (Zhu et al., 2019).
A preliminary study (Han et al., 2017) concluded that the sample of bones from this locality might contain two types of sauropod, and several species of theropod and hadrosaur. These bones are disarticulated and appear to have been rapidly buried in the course of a flooding event following transport over a short distance, as the clasts in the conglomerate are poorly rounded. Sauropod bones are particularly abundant in the sample, and Han et al. (2017) identified derived features suggesting that some of these bones were of titanosauriform origin whereas at least one, the left ischium ZMW148, was potentially from a diplodocoid. Relatively complete sauropod bones are known from the third and eighth units in the middle part of the section. The third unit is composed of siltstone and mainly contains cervical fragments, whereas the eighth is composed of conglomerate and is even richer in sauropod fossils, having produced the complete fibula, humerus, and ischium described here in addition to many unidentified fragments (Fig. 2).
Here, we present the first detailed study of the anatomically informative sauropod bones from the Tianyuan site, and reconsider their probable affinities within Sauropoda. The bones in the sample are not all proportionate in size to one another (see Tables 1–3) and in any case came from multiple stratigraphic levels, so multiple individuals are clearly represented. The bones may represent either a single species or multiple species.
| Elements | Dimension | Measurements |
|---|---|---|
| Cervical vertebra ZGT002 | Preserved centrum length (including ball) | 159 |
| Preserved centrum length (excluding ball) | 106 | |
| Anterior centrum height | 62.0 | |
| Anterior centrum width | 122 | |
| Preserved posterior centrum height | 77.7 | |
| Preserved posterior centrum width (dorsal) | 64.5 | |
| Preserved posterior centrum width (mid shaft) | 32.7 | |
| Neural arch height anteriorly | 58.5 | |
| Neural arch width anteriorly | 33.6 | |
| Width across prezygapophyses | 85.7 | |
| Anterior pneumatopore length | 31.6 | |
| Anterior pneumatopore height | 22.4 | |
| Posterior pneumatopore length | 67.7 | |
| Septum to posterior margin | 105 | |
| Posterior pneumatopore height (anterior) | 21.3 | |
| Anterior condyle width | 55.7 | |
| Anterior condyle height | 98.6 | |
| Preserved cervical rib length | 105 | |
| Caudal vertebra ZGT003 | Centrum length | 143 |
| anterior centrum height (without chevron facets) | 106 | |
| anterior centrum width (ventral/mid/dorsal regions) | 103/106/70 | |
| posterior centrum height (without chevron facets) | 106 | |
| posterior centrum width (ventral/mid/dorsal regions) | 92/99.6/76.8 | |
| Centrum height at the mid region | 96.2 | |
| Centrum width at the mid region | 82.6 | |
| Neural arch length (shortest) | 67.9 | |
| Neural arch width (mid region) | 55.0 | |
| Neural arch height | 43.4 | |
| Neural canal width/height (proximal end) | 35.0/38.4 | |
| Neural canal width/height (distal end) | 19.9/36.4 |
| Elements | Dimension | Measurements |
|---|---|---|
| Right humerus ZGT56-60 | Preserved length (medial/lateral margin) | 552/470 |
| Proximal end maximum mediolateral width | 345 | |
| Proximal end maximum anteroposterior thickness | 92.9 | |
| Distance from proximal end to distal edge of dtp crest | 420 | |
| Preserved mimimum shaft circumference | 410 | |
| Mediolateral width at midshaft | 175 | |
| Anteroposterior width at midshaft | 43.95 | |
| Dtp crest width (largest)/humerus width | 94.0/215 | |
| Left humerus ZGT089 | Preserved length | 318 |
| Largest transverse width as preserved | 151 | |
| Distance from proximal end to distal edge of dtp crest | 260 | |
| Mediolateral width at midshaft | 68.3 | |
| Anteroposterior width at midshaft | 48.9 | |
| Dtp crest width (largest)/humerus width | 56.7/151 |
Materials & Methods
All specimens described here are postcranial bones, including the following elements: ZGT002, a partial cervical vertebra lacking the posterior part; ZGT012, a partial cervical with the left rib preserved in articulation; ZGT005, two articulated, very incomplete cervical centra; ZGT044 and ZGT013, partial cervical ribs; ZGT003, a nearly complete caudal vertebra lacking the neural spine; ZMW143, a partial scapula; ZGT056-060, the well-preserved proximal half of a humerus; ZGT089, a midshaft fragment of a small humerus; ZMW148, a complete left ischium; ZMW51-57, a nearly complete fibula; and ZMW013, an almost complete pedal ungual. Some of these specimens are housed in the Bureau of Land and Resources of Zhuzhou City (ZGT), while others are in the Zhuzhou Museum (ZMW). Measurements of all these bones are presented in Tables 1–3. Comparisons with other taxa were mainly based on published literature and photographs. High-resolution 3D models of the cervical vertebra ZGT002 and the left ischium ZMW 148 are available in morphosourse.org, at https://www.morphosource.org/Detail/MediaDetail/Show/media_id/51457 for ZGT002 and https://www.morphosource.org/Detail/MediaDetail/Show/media_id/51458 for ZMW148. These bones were scanned using an Artec Space Spider hand-held 3D Scanner, and the scan data were edited to produce the final 3D models using Artec Studio.
Definitions of clades used in this study follow Mannion et al. (2013).
Results
Systematic paleontology
| Saurischia Seeley, 1887 |
| Sauropoda Marsh, 1878 |
| Neosauropoda Bonaparte, 1996 |
| Macronaria Wilson & Sereno, 1998 |
| Titanosauriformes Salgado, Coria and Calvo, 1997 |
| Titanosauriformes indet |
Description and comparisons
Cervical vertebrae and ribs
All of the cervical vertebrae described here were excavated from layer 3, in the middle part of the exposed section (Fig. 2). Two partial cervical vertebrae (ZGT002 and ZGT012), both with associated ribs, were identified (Figs. 3 and 4). ZGT002 is the better-preserved of the two and also exhibits some distinctive features. A detailed description of ZGT002, supplemented in places by information from the much less complete ZGT012, follows below. Measurements of ZGT002 are given in Table 1.
| Elements | Dimension | Measurements |
|---|---|---|
| Left ischium ZMW148 | Length | 595 |
| Anteroposterior length of iliac peduncle | 79.2 | |
| Maximum mediolateral width of iliac peduncle | 90.7 | |
| Dorsoventral height of iliac peduncle | 167 | |
| Anteroposterior length of acetabulum | 76.8 | |
| Anteroposterior length of proximal plate | 153 | |
| Dorsoventral height of pubic peduncle | 340 | |
| Maximum dorsoventral height of distal blade | 225 | |
| Maximum mediolateral width of distal blade | 31.4 | |
| Maximum anteroposterior length of distal blade | 60.2 | |
| Fibula ZMW51-57 | Length | 736 |
| Length from proximal end to proximal/distal end of lateral trochanter | 250/330 | |
| Mediolateral width of proximal end | 43.3 | |
| Anteroposterior length of proximal end | 225 | |
| Mediolateral width at midshaft | 32.2 | |
| Anteroposterior width at midshaft | 92.6 | |
| Minimum shaft circumference | 228 | |
| Mediolateral width of distal end | 75.6 | |
| Anteroposterior width of distal end | 129 | |
| Pedal ungual ZMW013 | Maximum proximodistal length | 137 |
| Proximal end dorsoventral height | 83.5 | |
| Proximal end mediolateral width | 47.9 |
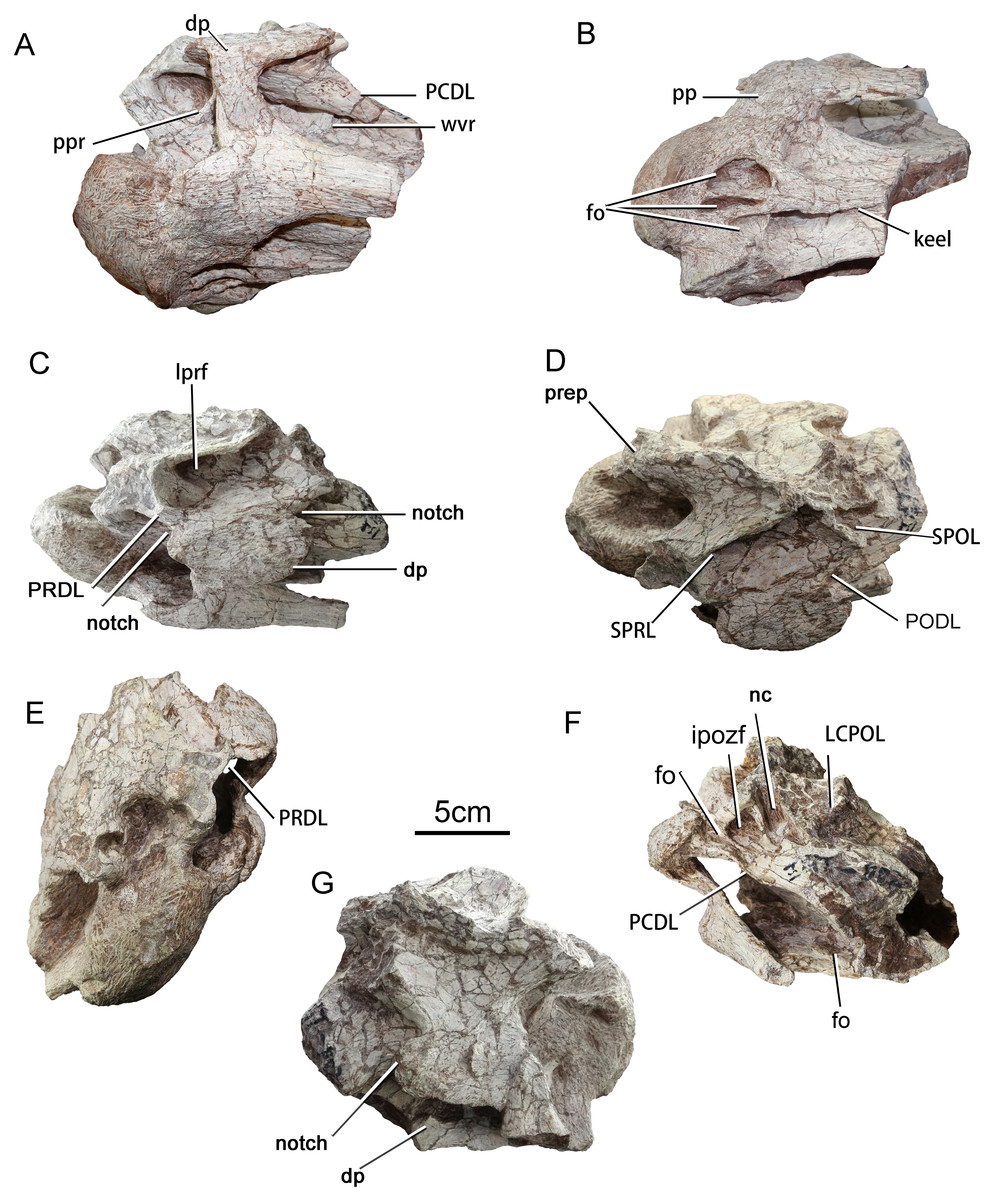
Figure 3: Cervical vertebra (ZGT002) of sauropod dinosaur from Zhuzhou City.
(A) Ventrolateral view. (B) Ventral view. (C) Left dorsolateral view. (D) Dorsal view. (E) Anterior view. (F) Posterior view. (G) Right lateral view. Abbreviations: dp, diapophysis; fo, fossa; ipozf, infrapostzygapophyseal fossa; LCPOL, lateral centropostzygapophyseal lamina; lprf, lateral fossa of the prezygapophyseal process; nc, neural canal; PCDL, posterior centrodiapophyseal lamina; PODL, postzygodiapophyseal lamina; pp, parapophysis; ppr, prominent posterodorsally oriented ridge; PRDL, prezygodiapophyseal lamina; SPRL, spinoprezygapophyseal lamina; SPOL, spinopostzygapophyseal lamina; wvr, weak vertically oriented ridge.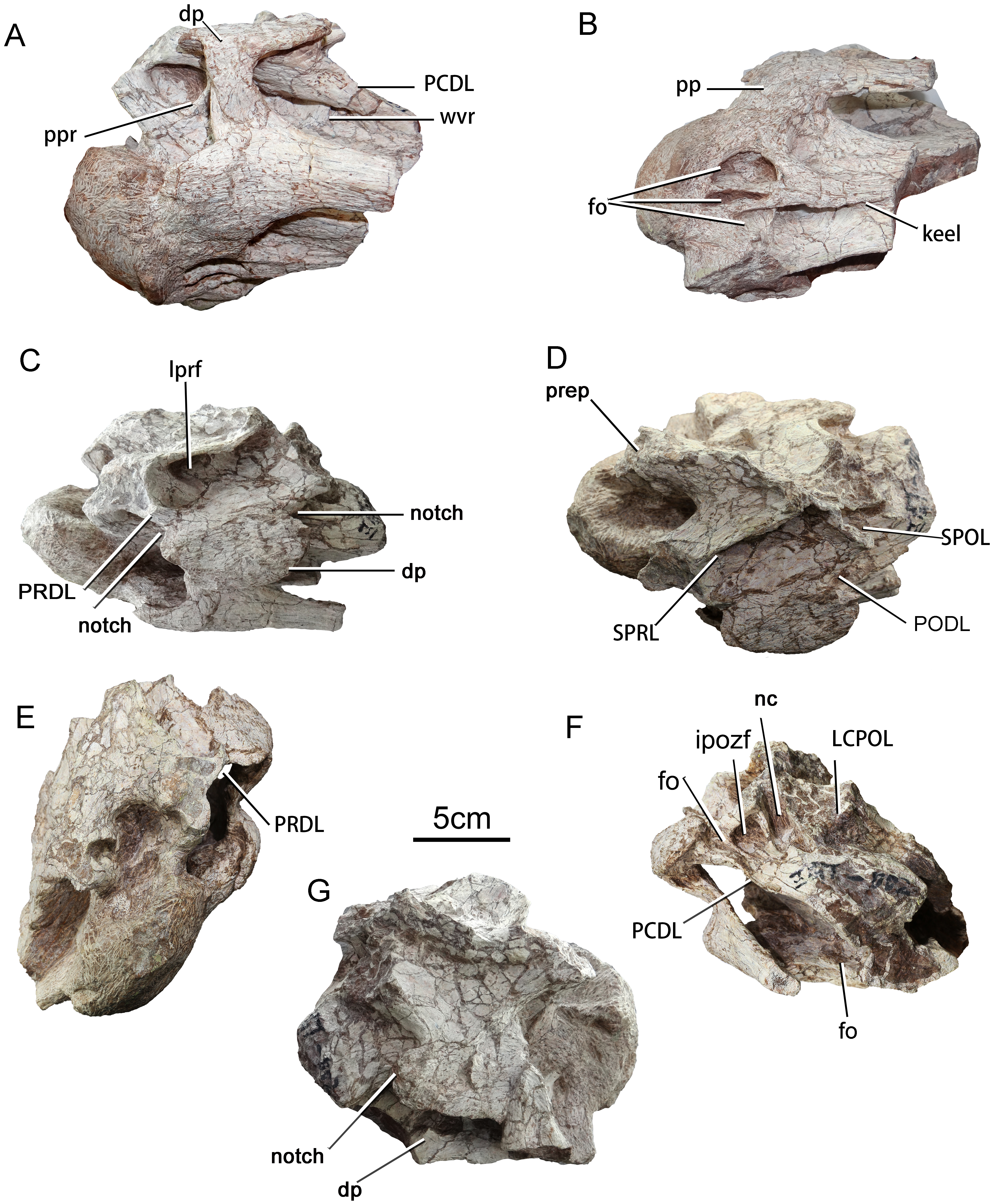{kind=link}

Figure 4: Sauropod dinosaur cervical vertebra (ZGT012) and ribs from Zhuzhou City.
(A–B) ZGT012, (A) centrum and associated rib in left lateral view; (B) vertebra without rib in left lateral view and detached rib in medial view. (C–E) ZGT044, large posterior cervical rib. (C) Dorsal view. (D) Posterior view. (E) Ventral view. (F–H) ZGT013, small posterior cervical rib. (F) Dorsal view. (G) Ventral view. (H) Medial view. Abbreviations: fo, fossa; ppr, prominent posterodorsally oriented ridge; rid, ridge.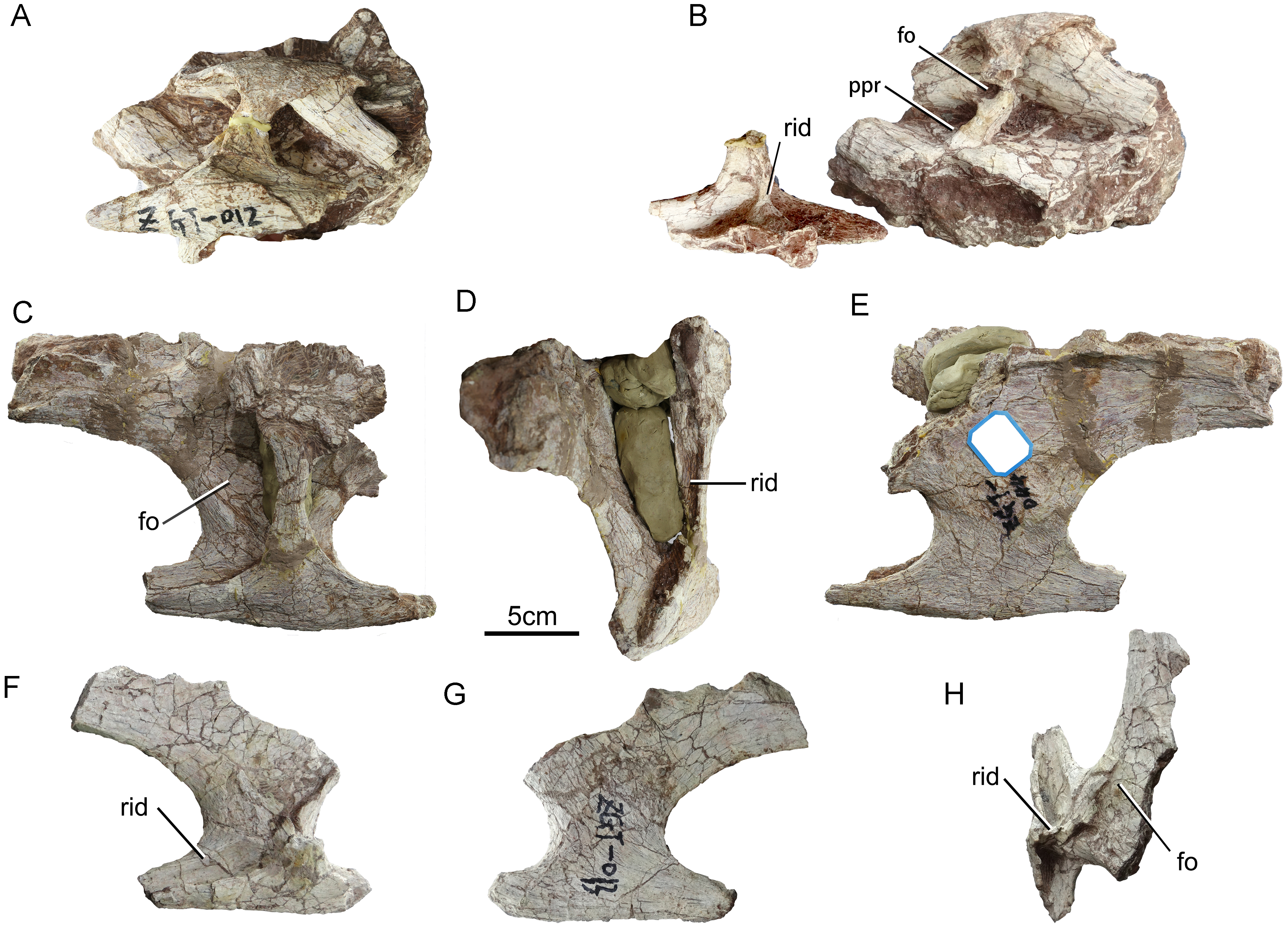{kind=link}
The anterior part of the centrum is well-preserved, but has undergone strong diagenetic compression in the dorsoventral direction. The portion of the centrum posterior to the diapophysis is broken away. The anterior articular surface is strongly convex, with a subcircular outline. The position of this partial vertebra within the cervical series is hard to assess with certainty, but on balance the specimen seems most likely to be from the anterior part of the neck. The most prominent part of the anterior condyle is displaced dorsally from the center of the articular surface as in the anterior and middle cervicals of the basal (non-titanosaurian) titanosauriform Euhelopus (Wilson & Upchurch, 2009), although in the anterior cervicals of Erketu and Yunmenglong the convexity of the anterior condyle is more uniform (Ksepka & Norell, 2006; Lü et al., 2013a). The lateral surface of the centrum is slightly concave, which are more consistent with attribution to the anterior part of the cervical column, as posterior cervicals tend to bear more elaborate laminae and pneumatic cavities than anterior ones (Gomani, 2005). The neural spine is missing, but the cross section of the broken base of the spine is thin and short, suggesting that the intact spine would have been of the poorly developed type expected in an anterior cervical. In Euhelopus the neural spines become transversely wide and bifurcated posterior to cervical 11 (Wilson & Upchurch, 2009). If ZGT002 is indeed an anterior cervical vertebra, comparison with the anterior cervicals of Erketu (Ksepka & Norell, 2006) suggests that more than half of the centrum is missing.
The left lateral surface of the centrum is depressed, and the ventral part of this surface is further excavated by an anteroposteriorly elongate fossa that can be clearly seen in posterior view (Fig. 3F) and also in the 3D model. The anterior cervical centra of Erketu display a similar condition, albeit with a deeper fossa (Ksepka & Norell, 2006). In comparison, two or three pairs of small deep pleurocoels occur in the anterior cervicals of Yunmenglong (Lü et al., 2013a) and Qiaowanlong (You & Li, 2009), whereas in the preserved posterior cervical vertebrae of the titanosauriforms Daxiatitan and Yongjinglong only a single large, deep fossa is present on each lateral surface of the centrum (You et al., 2008; Li et al., 2014).
In ventrolateral view, it can be clearly seen that the neural arch is excavated by two infradiapophyseal fossae separated by a prominent, posterodorsally oriented ridge (ppr, Fig. 3A). This ridge is more robust in ZGT012 than in ZGT002 (Fig. 4B). In ZGT012, a small, deep fossa penetrates the widened posterodorsal end of the lamina (fo, Fig. 4B). The anterior infradiapophyseal fossa is subtriangular in lateral view, whereas the posterior one is elongate and shallow dorsoventrally. In ZGT002, a weak, vertically oriented secondary ridge separates the posterior fossa into an anterior subrectangular compartment and a posterior shallow triangular compartment (Fig. 3A).
In ZGT002 the diapophysis arises from the dorsal portion of the centrum and has been deflected upward as a result of distortion (Figs. 3C and 3G). A narrow process extends ventrolaterally from near the anterior end of the anteroposteriorly elongate base of the diapophysis, so that the diapophysis as a whole has a rotated “L” shape in lateral view. On the left side of the vertebra, the base of the diapophysis is embayed by small notches both anteriorly and posteriorly (Fig. 3C). On the right side, however, only a posterior notch is present, suggesting that the anterior notch on the left diapophysis is a result of damage (Fig. 3G). The ventral process of the diapophysis overlaps the dorsolateral surface of the tuberculum. The long, shallow parapophysis extends laterally from the anteroventral corner of the centrum and is fused with the capitulum of the cervical rib. In other titanosauriforms such as Euhelopus, Erketu and Huabeisaurus (Ksepka & Norell, 2006; Wilson & Upchurch, 2009; D’Emic et al., 2013), the parapophysis has a strong ventrolateral inclination, and the lateral orientation of the parapophysis of ZGT002 may be a result of taphonomic deformation.
The anterior part of the ventral surface of the centrum bears three adjacent large fossae, whereas the middle part bears a prominent midline keel (Figs. 3A and 3B). In many titanosauriforms (e.g., Qiaowanlong You & Li, 2009; Euhelopus Wilson & Upchurch, 2009), the ventral surface is concave and a keel is lacking. The right fossa is semicircular in outline and is relatively small and shallow, whereas the midline fossa has an oblong outline and is longer, narrower and deeper. The left fossa is similar in length to the midline fossa but is much wider, and semicircular in outline. The three fossae are separated by long, sharp ridges. A shallow depression is situated on the ventral surface of the midline keel, just posterior to the medial part of the left fossa, and is separated from the left fossa by a short but prominent transverse ridge. The shapes and sizes of the three fossae may be affected by the overall distortion of the vertebra. Cervical vertebrae with paired fossae in the same position as the three fossae seen in ZGT002 are known in some titanosaurs, including Rukwatitan (posterior cervical, Gorscak et al., 2014) and Overosaurus (posterior cervical, Coria et al., 2013). Paired fossae have also been reported in the cervicals of Lingwulong (Xu et al., 2018) and some other dicraeosaurids (Whitlock, D’Emic & Wilson, 2011), but these are separated by a keel (Tschopp, Mateus & Benson, 2015: Fig. 38) whereas in ZGT002 the ventral keel begins posterior to the fossae. In titanosaurs with paired fossae, the presence of a keel separating the fossae is variable (Gorscak et al., 2014).
The presence of a trio of fossae at the anterior end of the ventral surface of the centrum of ZGT002 is unusual, having never been reported in another sauropod cervical vertebra, and may potentially represent an autapomorphy indicating that at least some of the Zhuzhou sauropod material can be referred to a new species. However, we refrain from erecting a new species pending recovery of more complete specimens, particularly given the paucity of other clear potential autapomorphies in the material and the fact that pneumatic features such as fossae can be highly variable.
The keel occupying the middle part of the ventral surface of the centrum is transversely wide anteriorly and narrows posteriorly, taking on a wedge-like shape. The keel essentially divides the middle part of the ventral surface into two large, shallow depressions (Fig. 3B). A ventral keel is a primitive character that appears in the cervicals of some non-neosauropods (e.g., Shunosaurus) and some diplodocoids, occurring in most cervicals of Dicraeosaurus (Harris, 2006: Fig. 2), the middle and posterior cervicals of Kaatedocus (Tschopp & Mateus, 2013) and the only known reasonably intact anterior cervical of Katepensaurus (Ibiricu et al., 2013: Fig. 3C). A ventral keel is also seen in some titanosauriform cervicals, such as the axis of Erketu (Ksepka & Norell, 2006: Fig. 3), the axis and anterior cervicals of Mongolosaurus (Mannion, 2011: Fig. 6), the axis of Futalognkosaurus (Mannion, 2011), the only known (posterior) cervical of Savannasaurus (Poropat et al., 2016), and the cervicals of Rapetosaurus (Curry Rogers, 2009). A weak midline ridge is also present on the anterior part of the ventral surface of the centrum in cervicals 3 and 17 of Euhelopus (Wilson & Upchurch, 2009). Taken together, these observations imply that ventral keels can occur throughout the cervical series in non-neosauropods, diplodocoids and titanosauriforms. The ventral keel seen in ZGT002 resembles those on the axial vertebrae of Erketu and Mongolosaurus (Ksepka & Norell, 2006; Mannion, 2011) in being transversely thickened anteriorly and becoming narrower and shallower posteriorly, providing more evidence that ZGT002 may represent an axis or anterior postaxial cervical.
Loss through breakage of the posterior portion of the centrum has resulted in the exposure of internal cavities. In posterior view, the interior of the centrum can be seen to be divided into two large chambers (camerae) by a longitudinal lamina (Fig. 3F), as in basal sauropods such as Camarasaurus (Wedel, Cifelli & Sanders, 2000). However, the interior of the neural arch region has a porous appearance, being divided irregularly into smaller chambers (camellae) as in titanosauriforms such as Euhelopus (Wilson & Upchurch, 2009). The combination of a largely camerate centrum and an at least partly camellate neural arch is evident in basal titanosauriforms, such as Brachiosaurus (Wedel, Cifelli & Sanders, 2000: Fig. 12C, BYU12866), but differs from the fully camellate condition typically seen in derived titanosaurians, such as Saltasaurus (Wedel, 2003).
There is no visible suture at the neurocentral junction, and the neural arch is damaged, although the neural canal is large. The pre- and postzygapophyses are missing. The PRDL (prezygodiapophyseal lamina) is robust and extends posteroventrally from the lateral side of the prezygapophysis to the base of the diapophysis (Fig. 3C). Most of the neural spine is missing, but the preserved basal part is transversely thin, suggesting that this structure was non-bifid. In contrast, bifurcated neural spines with transversely thick bases are present in some of the presacral vertebrae of many sauropods (Wedel & Taylor, 2013), including Euhelopus (Wilson & Upchurch, 2009) and Erketu (Ksepka & Norell, 2006). The paired spinoprezygapophyseal laminae are well-preserved, and are thin and deep with sharp edges. These laminae diverge anterolaterally from the spine, forming between them an angle of 45°, and merge into the prezygapophyses. There is no prespinal lamina or fossa.
In posterior view, it is evident that the centrum and neural arch are both distorted. The neural canal is subrectangular, being dorsoventrally elongate and transversely narrow. The left infrapostzygapophyseal fossa (ipozf, Fig. 3F) is well preserved. This fossa is subtriangular in outline and bounded dorsolaterally by the postzygodiapophyseal lamina (PODL), ventrolaterally by a laterally curved lamina, and medially by a vertical lamina that separates the fossa from the neural canal. A second shallow fossa (fo, Fig. 3F) is situated just lateral to the large infrapostzygopophyseal fossa. The small fossa is dorsoventrally elongate but transversely narrow.
The anterior part of the left cervical rib belonging to ZGT002 is well-preserved, but the posterior part is broken away. The rib shaft is robust and straight, and narrows dorsoventrally towards the missing posterior end. The lateral surface is strongly dorsoventrally convex whereas the medial surface is flatter, producing a semi-circular cross-section, as in the anterior to mid cervical ribs of Kaatedocus (Tschopp & Mateus, 2013). The anterior part is divided into a slender dorsal tuberculum and a deep, short ventral capitulum. The capitulum is fused to the parapophysis of ZGT002 as described above, but the tuberculum is not fused to the diapophysis (Fig. 3A). Three other cervical ribs are also well-preserved. In ZGT012, the tuberculum is fused to the diapophysis, and the parapophysis and capitulum are broken away. The angle between the tuberculum and capitulum is about 90°, as in basal sauropods such as Shunosaurus (Dong, Zhou & Zhang, 1983), but may have been altered by preservational distortion. The cervical rib bears a long anterior process, which is transversely narrow, tapers gradually to a point, and has a convex lateral surface and a concave medial one. On the medial surface of the rib (Fig. 4B), a ridge extends upwards from the base of the capitulum along the anterior margin of the tuberculum. Most of the shaft is missing, but the base of the shaft is dorsoventrally deep and transversely narrow. The capitulum is broken through near its medial end, and the cross-section shows several small cavities.
Cervical ribs ZGT044 and ZGT013 are similar to each other but differ in size (Fig. 4). The positions of these ribs within the cervical series are unknown. ZGT044 is double-headed, and fused at the capitulum and tuberculum to a fragment of the corresponding vertebra. In ZGT013, the tuberculum is mostly broken away, whereas the capitulum is fused to a fragment of the elongate parapophysis of the corresponding vertebra. A moderately well-developed ridge extends posteromedially across the dorsal surface of the base of the posterior process (Fig. 4F). In both these ribs, the angle between the capitulum and the tuberculum is about 45°, as in Omeisaurus and neosauropods (Wilson & Sereno, 1998). The tuberculum is anteroposteriorly narrow, and has a thin anterior margin but a thick posterior one. The capitulum is wide anteroposteriorly and thin dorsoventrally. The dorsal surface of the capitulum bears a large, shallow fossa (Figs. 4C and 4H), as in some basal sauropods (Upchurch, Barrett & Dodson, 2004). The medial surface of the tuberculum is visible in ZGT012, ZGT013, ZGT044. A robust vertical ridge emanates from the midpoint of the base of the tuberculum and extends dorsally across the entire medial face, curving posteriorly near its dorsal end (Figs. 4B, 4D and 4H).
The anterior and posterior processes of both ZGT044 and ZGT013 are well-developed. In both cases the anterior process is slightly damaged at the tapering, anterodorsally curving tip, whereas the posterior process is only partially preserved.
Caudal vertebra (ZGT003)
A single, almost complete caudal vertebra (ZGT003) is present (Fig. 5). This specimen appears to represent a middle caudal vertebra, based on its large size, elongate proportions and lack of transverse processes. The centrum is 143 mm long, 100 mm wide, and 109 mm tall (Table 1). By contrast, most sauropod anterior caudal centra, including those of the first eleven caudals of Tambatitanis (Saegusa & Ikeda, 2014) and the anterior 10 caudals of Huanghetitan ruyangensis (Lü et al., 2007), are wider than long. The length of the centra tends to increase posteriorly along the caudal series (Upchurch, Barrett & Dodson, 2004), so that centrum length and centrum width are usually about equal in caudals 10–15 (You, Tang & Luo, 2003). Therefore, ZGT003 is probably from the part of the tail posterior to caudal 10.

Figure 5: Caudal vertebra of sauropod dinosaur from Zhuzhou City (ZGT013).
A, left lateral view; B, right lateral view; C, anterior view; D, posterior view; E, dorsal view; F, ventral view. Abbreviation: rid, ridge.{kind=link}
The centrum is slightly amphicoelous and subcylindrical, as in the middle caudals of basal titanosauriforms such as Gobititan (You, Tang & Luo, 2003) and Phuwiangosaurus (Martin, 1999: Figs. 25 and 26). In Huanghetitan ruyangensis, the anteriormost five caudal centra are amphicoelous, whereas those of caudals 6 to 10 are amphiplatyan (Lü et al., 2007). In contrast, the anterior and middle caudals are procoelous in some titanosauriforms, including Daxiatitan (You et al., 2008) and Tambatitanis (Saegusa & Ikeda, 2014). In Mamenchisaurus, the 15 anteriormost caudal centra are procoelous, whereas the centra are amphiplatyan from caudal 16 onward (Young & Zhao, 1972). In Borealosaurus (You et al., 2004) and Fukuititan (Azuma & Shibata, 2010), the only preserved caudal centra are middle or distal in position, and are opisthocoelous.
The lateral surface of the centrum is strongly dorsoventrally convex, and bears a weak longitudinal ridge at about two-thirds of its height (Fig. 5B). The presence of the ridge gives the cross-section of the centrum a hexagonal outline, as in the eighth and ninth caudal vertebrae of Huanghetitan (Lü et al., 2007) and middle caudal vertebrae of the basal titanosaur Andesaurus (Mannion & Calvo, 2011: Fig. 6). Several nutrient foramina are present on the lateral surfaces of the centrum, as in Ganannosaurus (Lü et al., 2013b).
The anterior and posterior articular surfaces are both well-preserved and expanded dorsoventrally, the anterior surface being the larger of the two. In anterior or posterior view, the outline of the centrum resembles a hexagon with slightly convex sides and a slightly concave top and bottom, the hexagon achieving its greatest transverse width at a level below its dorsoventral midpoint. The ventral surface of the centrum is smoothly concave, and terminates anteriorly at a transverse ridge (Fig. 5F). The apex of the concavity lies just below the posterior part of the neural arch, and the ventral surface bears weak longitudinal ridges near its lateral margins (Fig. 5A). However, the ventral surface lacks a midline groove, a feature present in many forms (Upchurch, Barrett & Dodson, 2004) including Huanghetitan (caudals 8 and 9, Lü et al., 2007). The ventral surface also lacks the anteroposterior row of distinct fossae that occur in the anterior caudals of derived diplodocids and lithostrotians (Upchurch, Barrett & Dodson, 2004).
The anterior and posterior pairs of chevron facets are both well-developed. The anterior facets are convex, directed ventrally, and subcircular in outline. The posterior facets are relatively flattened, directed more posteriorly than ventrally, and subtriangular in outline.
The neural canal is very small compared to the centrum, being only about 1/5 as high as the latter. The width and height of the anterior opening are approximately equal, whereas the posterior opening is only two thirds as wide as high. The neural arch is restricted to the anterior half of the centrum as in the middle caudal vertebrae of Phuwiangosaurus (Martin, 1999: Fig. 26) and most other titanosauriforms (Upchurch, 1998). In Gobititan the neural arch is placed near the mid-length of the centrum in the mid-caudals, but is entirely restricted to the anterior part of the centrum in caudal 33 (You, Tang & Luo, 2003). The neural spine and zygapophyses are broken away, although the base of the left prezygapophysis is partially preserved. The prezygapophysis appears to have originally extended anterodorsally, and has a transversely convex ventral surface. The damaged surfaces of the neural arch are honeycombed with pneumatic chambers in a camellate pattern, as in an anterior caudal of the titanosaurian Malawisaurus (Wedel, 2009).
Humerus
Two well-preserved partial humeri of different sizes (ZGT56-60 and ZGT089) are present in the sample (Fig. 6). ZGT56-60 is the larger and more completely preserved of the two. Measurements for these two humeri are given in Table 2.

Figure 6: Sauropod dinosaur humeri from Zhuzhou City.
(A–E) ZGT056-060, partial right humerus. (A) Anterior view; (B) lateral view; (C) posterior view; (D) proximal view; (E) distal view of broken surface. (F–H) ZGT089, partial left humerus. (F) Distal view of broken surface; (G) anterior view; (H) posterior view. Abbreviations: dp, deltopectoral crest; hh, humeral head, lbh, lateral bulge of humerus; hplc, humeral proximolateral corner.{kind=link}
ZGT56-60 is the proximal half of a right humerus, and is slightly compressed anteroposteriorly. The long axis of this partial humerus appears straight in lateral view, and the proximal end is transversely wide. Although the distal end is absent, the preserved portion of the humerus extends more or less to the midshaft region, and indicates that the humerus as a whole was robust.
The proximal end of the humerus is extremely expanded transversely, its width of 40 cm being about double that of the distalmost preserved part of the humeral shaft. The head of the humerus is proximally convex and forms a strong medial prominence as in other sauropods (Upchurch, Barrett & Dodson, 2004). The proximolateral corner is relatively rounded, and subdued relative to the proximal apex of the humeral head, as in the basal titanosauriform Qingxiusaurus (Mo et al., 2008: Fig. 1). By contrast, the proximolateral corner has a well-defined square outline in most somphospondylans (Upchurch, Barrett & Dodson, 2004). The medial part of the proximal articular surface is slightly convex in anterior view, and the articular surface as a whole does not show the sigmoid curvature that is present in some titanosaurians, such as Diamantinasaurus (Poropat et al., 2015) and Opisthocoelicaudia (Borsuk-Białynicka, 1977). The proximal articular surface is rugose, crescent-shaped, and unevenly expanded anteroposteriorly to form the humeral head, the greatest expansion occurring around the mediolateral midpoint of the proximal surface. This is unlike the anteroposteriorly narrow humeral head in Euhelopus (Young, 1935; Wilson & Upchurch, 2009). The medial end of the proximal articular surface tapers gradually to a point, whereas approximately the lateralmost third of the proximal surface has a subrectangular outline, is distinctly narrower than the expanded humeral head, and slopes down gently towards the deltopectoral crest. No fossae or tubercles interrupt the proximal articular surface. The convex humeral head is only moderately well-developed and merges gradually into the rest of the proximal end of the humerus. In some macronarian taxa, such as Haestasaurus (Upchurch, Mannion & Taylor, 2015), the humeral head is more prominent and sharply defined. Well distal to the proximal articular surface, a bulge is present on the lateral side of the humerus posterior to the deltopectoral crest. A scar or eminence in this area occurs in a variety of titanosauriforms and is conventionally identified as marking the insertion of M. scapulohumeralis anterior (Borsuk-Białynicka, 1977; Upchurch, Mannion & Taylor, 2015). The bulge is prominent and clearly visible in anterior view, as in taxa such as Qingxiusaurus (Mo et al., 2008) and Neuquensaurus (Otero, 2010).
The deltopectoral crest is situated along the anterolateral margin of the humerus and extends to the distal part of the preserved portion. The crest is low, straight, and thickened mediolaterally in its midlength portion. This is unlike the strongly medially directed deltopectoral crest seen in the titanosaurs Opisthocoelicaudia (Borsuk-Białynicka, 1977) and Zhuchengtitan (Mo et al., 2017). The deltopectoral crest is a subdued structure with a flattened anterior surface, unlike the more prominent and ridge-like deltopectoral crest of Qingxiusaurus (Mo et al., 2008: Fig. 1). In anterior or posterior view the lateral margin of the shaft appears relatively straight in ZGT56-60 but concave in the small humerus ZGT089, a variation that may reflect ontogenetic change. A straight lateral humeral margin has been regarded as a derived feature only seen in some titanosauriforms, such as Malawisaurus (Gomani, 2005).
The proximal part of the cranial surface is mediolaterally concave, and is cracked due to poor preservation. No rugosities are apparent on this part of the humeral surface. The posterior surface is convex, giving the cross-section of this part of the humerus a thin, crescentic shape. In ZGT56-60, the cross-section of the middle part of the shaft is three times as mediolaterally wide as anteroposteriorly thick, but in ZGT089 the cross-section is oval and only slightly wider than thick (Fig. 6). The extreme midshaft thinness of ZGT56-60 may be a result of the compression that caused the cracks on the anterior surface.
Ischium
The left ischium ZMW148 is well-preserved and nearly complete (Fig. 7), and is flattened, anteroposteriorly wide, and dorsoventrally elongate. The medial surface remains obscured by matrix. A 3D model has been prepared for detailed examination and comparison. The ischium has a dorsoventral length of 59.5 cm (Table 3) and is characterized by a short, robust iliac peduncle, a wide, elongate pubic peduncle, and a slender, rod-like posteroventral shaft. The maximum width of the middle portion of the ischium is more than twice that of the iliac peduncle or the distal shaft.
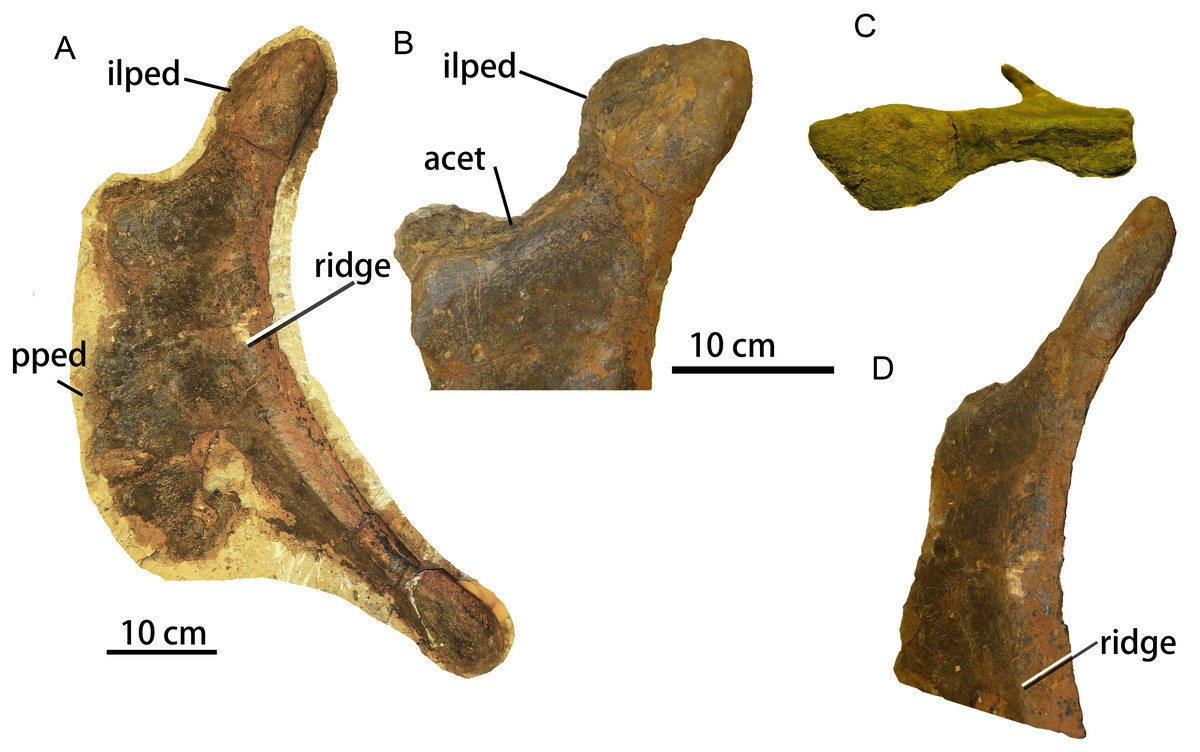
Figure 7: Complete left ischium of sauropod dinosaur from Zhuzhou City (ZMW148).
A, left lateral view; B, enlargement of the proximal region in lateral view; C, proximal region in dorsolateral view; D, proximal region in posterolateral view. Abbreviations: acet, acetabulum. pped, pubic peduncle; ilped, iliac peduncle.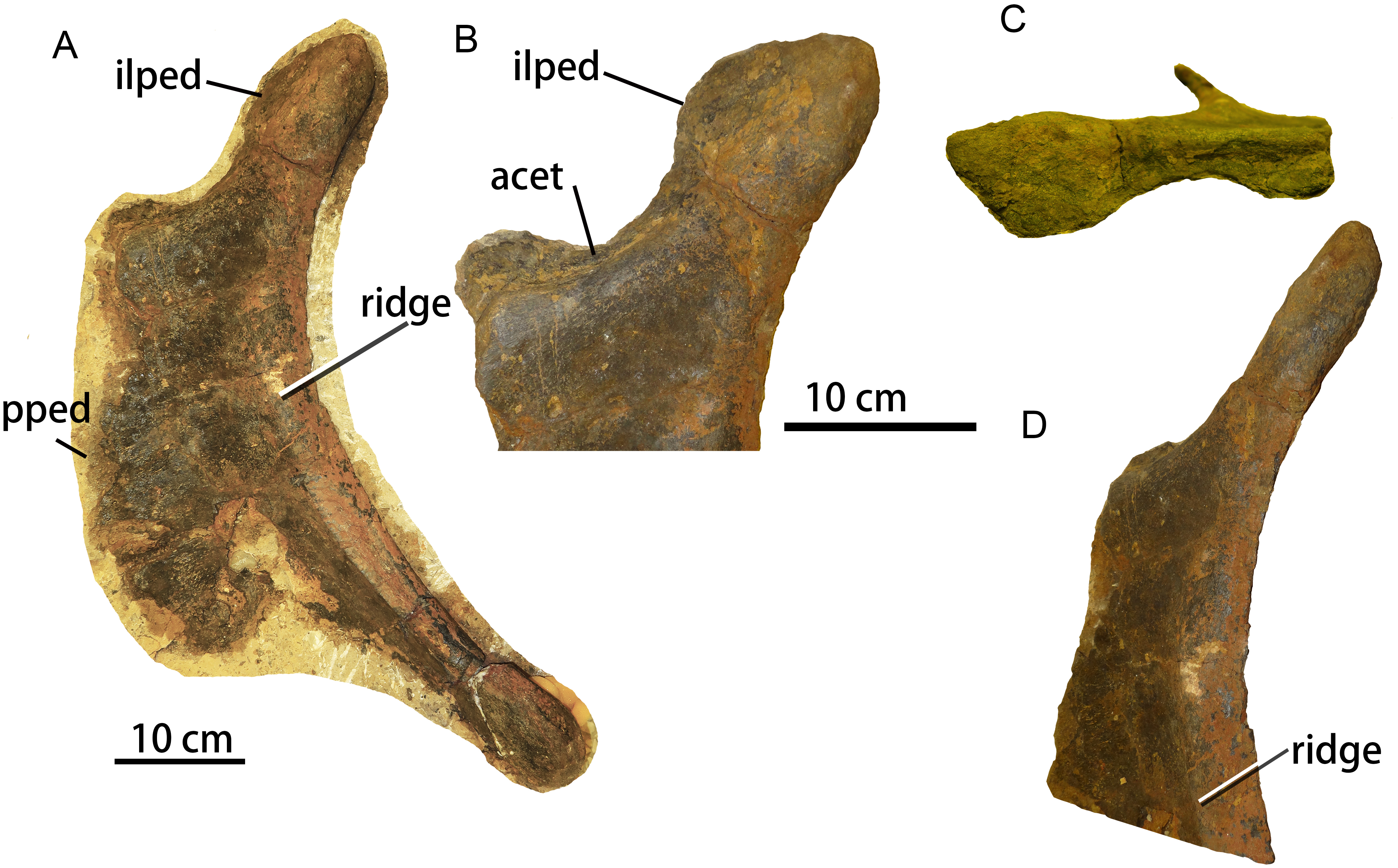{kind=link}
The iliac peduncle accounts for about one-fourth of the proximodistal length of the ischium, and contributes a large portion to the margin of the acetabulum. The articular surface for the ilium faces anterolaterally, while the iliac peduncle as a whole thickens posteriorly and therefore has a subtriangular cross-section. The anterior margin of the iliac peduncle is continuous with the dorsal margin of the pubic peduncle. The iliac peduncle has a relatively thin stalk but is transversely expanded at the dorsal end for the articulation with the ilium. The acetabular surface of the ischium is mediolaterally narrowest in its central portion (Fig. 7C), which occurs in most rebbachisaurids (Mannion et al., 2012) but has also been reported in the titanosauriform Huabeisaurus from the Upper Cretaceous of Shanxi Province, China (D’Emic et al., 2013: Fig. 20).
The concave posterior margin of the ischium is thickened, forming a prominent ridge that extends from the base of the iliac peduncle to the posteroventral end of the ischial shaft (Fig. 7A). This ridge is damaged in the mid-region of the posterior margin (see 3D model). A similar ridge is also seen in the brachiosaurid Soriatitan (Royo-Torres et al., 2017: Fig. 9). There is no ischial tuberosity on the lateral surface of the ischium, unlike the condition in Huabeisaurus (D’Emic et al., 2013) and Neuquensaurus australis (see Otero, 2010: Figs. 8A–8B).
The pubic peduncle is mediolaterally thin and has a flat lateral surface. Its anterior margin has an irregular appearance resulting from damage. The dorsoventral height of the pubic peduncle is about triple its anteroposterior length, as in the titanosauriforms Sonidosaurus (Xu et al., 2006) and Qiaowanlong (You & Li, 2009: Fig. 3). The ventral margin of the ischium is thin and somewhat damaged.
The length of the posteroventral shaft is slightly less than the dorsoventral height of the ischial peduncle. The posterior edge of the shaft is transversely expanded and rounded whereas the anterior edge is thin and sheet-like, so that the shaft has a subtriangular cross-section. The shaft is extremely narrow anteroposteriorly, in contrast to the wide shaft seen in most titanosauriforms, such as Huabeisaurus (D’Emic et al., 2013), Qiaowanlong (You & Li, 2009: Fig. 3), Sonidosaurus (Xu et al., 2006) and Rapetosaurus (Curry Rogers, 2009: Fig. 41). However, an ischium with a narrow distal shaft also occurs in some titanosauriforms, such as Dongyangsaurus (Lü et al., 2008, Plate II), Opisthocoelicaudia and an unnamed Brazilian taxon (Lehman & Coulson, 2002), as well as in most rebbachisaurids including Comahuesaurus (Carballido et al., 2012: Fig. 10), Zapalasaurus (Salgado, Carvalho & Garrido, 2006: Fig. 6) and Rebbachisaurus (Wilson & Allain, 2015: Fig. 15). The distal end of the shaft is unexpanded, as in the brachiosaurid Soriatitan (Royo-Torres et al., 2017) and such rebbachisaurids as Comahuesaurus, Zapalasaurus (Salgado, Carvalho & Garrido, 2006; Carballido et al., 2012). In most titanosauriforms, by contrast, including Qiaowanlong (You & Li, 2009) and Huabeisaurus (D’Emic et al., 2013), the distal end of the shaft is anteroposteriorly expanded. The anteroposterior length of the distal end is only twice the mediolateral width, compared to three times the width in Sonidosaurus (Xu et al., 2006) and five times the width in Huabeisaurus (D’Emic et al., 2013).
In general, the ischium is similar to those of other titanosauriforms, especially in having a dorsoventrally elongate pubic peduncle. However, the ischium is also unusual in having some features that are otherwise mainly known in rebbachisaurids, including a slender shaft and a narrow central portion of the acetabular surface.
Fibula
The long and slender right fibula ZMW 51-57 is well-preserved, except that part of the midshaft region is missing (Fig. 8). Measurements of the fibula are provided in Table 3. In lateral view, the anterior margin of the fibula is relatively straight, whereas the posterior margin is strongly concave. The proximal end is transversely compressed and flares asymmetrically in lateral view, being expanded posteriorly but not anteriorly. The proximal end has a crescentic outline, being convex laterally and concave medially, and the posterior part of the proximal end is thicker than the anterior part. Additionally, the proximal end lacks the anteromedial crest, which fits into a notch on the tibia, seen in such titanosauriforms as Gobititan (You, Tang & Luo, 2003: Fig. 2), Erketu (Ksepka & Norell, 2006: Fig. 10) and Euhelopus (Wilson & Upchurch, 2009: Fig. 25)
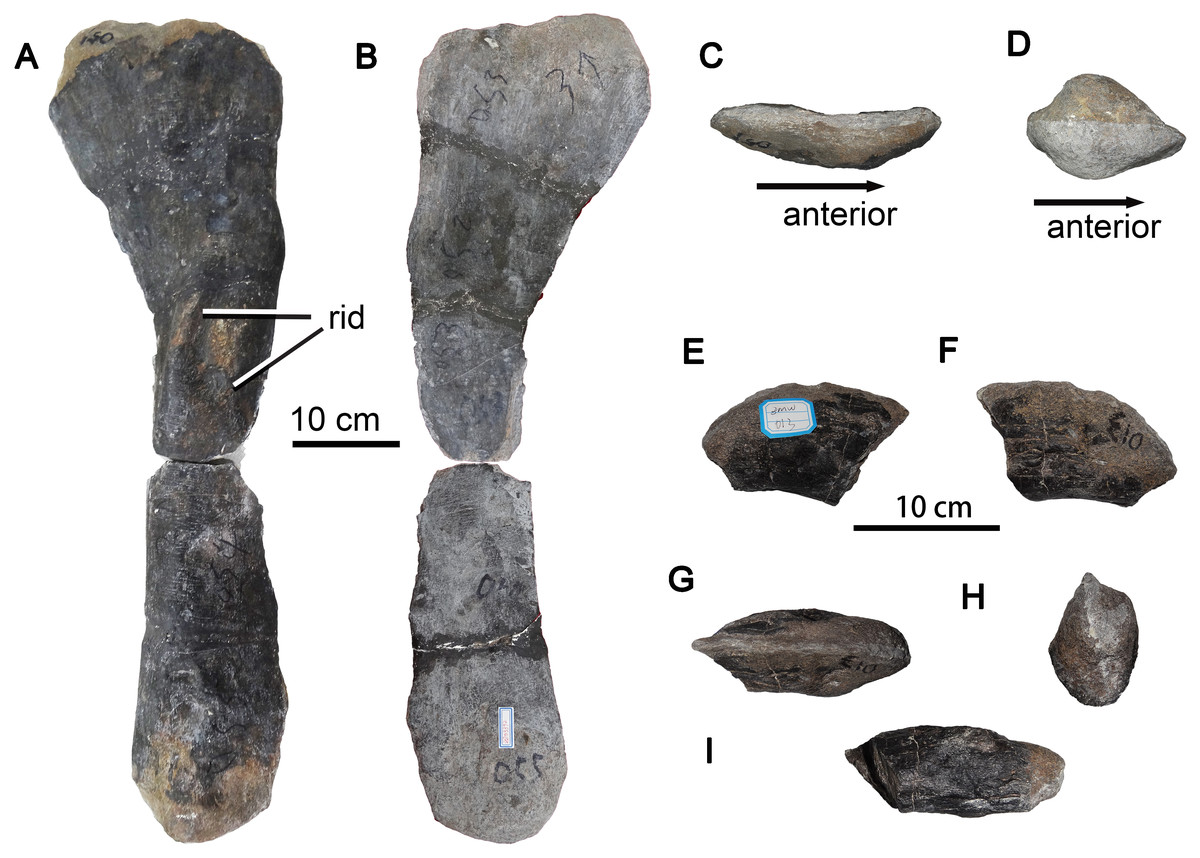
Figure 8: Sauropod dinosaur right fibula and ungual from Zhuzhou City.
(A–D) ZMW51-57, nearly complete right fibula. (A) Right lateral view; (B) medial view; (C) proximal view; (D) distal view. (E–G) ZMW013, a pedal ungual. (E) Left lateral view; (F) right lateral view; (G) dorsal view; (H) proximal view; (I) ventral view. Abbreviations: rid, ridge.{kind=link}
The anteroproximal portion of the fibula appears rounded in lateral view. The strong posterior expansion of the proximalmost part of the fibula has a subtriangular outline in lateral view, although the posterior apex is truncated (Fig. 8A). The lateral surface of the shaft remains strongly convex over the entire length of the fibula, whereas the medial surface is concave over the proximal half of the bone but slightly convex over the distal half.
A lateral trochanter is present near the midpoint of the shaft. The trochanter is slightly curved and anterodorsally oriented, and takes the form of two elongate, narrow parallel ridges as observed in many titanosauriforms, such as Huabeisaurus (D’Emic et al., 2013: Fig. 23). Deep fossae are present both anterior and posterior to the more posterior of the two ridges. The distal end of the fibula is expanded relative to the midshaft, strongly convex both anteroposteriorly and transversely, and lenticular in outline.
Ungual
A well-preserved, nearly complete large pedal ungual (ZMW013), with a length of about 10 cm, is present. This element is strongly compressed laterally, with slightly dorsoventrally convex lateral and medial sides, as in other Eusauropoda (Wilson & Sereno, 1998). The proximal end of the ungual is dorsoventrally deep, transversely narrow, and symmetrical in outline. The dorsal margin of the ungual is sharp and curved anteroventrally, whereas the ventral surface is transversely expanded and flattened (Fig. 8I). Which pedal digit the ungual belongs to is uncertain.
Discussion
Titanosauriforms are relatively diverse and abundant in the Cretaceous of eastern Asia, at least 20 species having been erected (D’Emic et al., 2013; Mo et al., 2017; Averianov & Skutschas, 2017; Averianov et al., 2018). Wilson (2005) noted that all Cretaceous sauropods from eastern Asia were titanosauriforms, whereas all Jurassic sauropods from the region were non-neosauropods, although this situation changed with the discovery of the Jurassic diplodocoid Lingwulong (Xu et al., 2018). The Zhuzhou sauropod specimens add to the wealth of fossil evidence regarding Cretaceous eastern Asian titanosauriforms, and collectively display an interesting and complex combination of morphological characters.
Many authors have suggested some degree of endemicity among eastern Asian sauropods in general, and have proposed the existence of putative endemic eastern Asian titanosauriform clades including Euhelopodidae (Wilson & Upchurch, 2009; D’Emic, 2012), Nemegtosauridae (Wilson, 2005) and Opisthocoelicaudiinae (You et al., 2004). Recent phylogenies support the validity of Euhelopodidae, but the validity of Nemegtosauridae and Opisthocoelicaudiinae remains controversial (Upchurch, Mannion & Taylor, 2015; Poropat et al., 2016; González Riga et al., 2018; Mannion et al., 2019). Arriving at a clearer understanding of titanosauriform phylogeny will undoubtedly require more extensive description of some of the lesser-studied Cretaceous eastern Asian taxa, ideally based in some cases on much more complete specimens than are presently available. In this connection, the Zhuzhou sauropod material is unfortunately so fragmentary that it is difficult to establish its phylogenetic position within Titanosauriformes, or even to determine whether more than one taxon might be present in the sample. Nevertheless, some if not all of the Zhuzhou sauropod bones are clearly of somphospondylan origin. Somphospondylans were among the most abundant and widespread sauropods, and were particularly diverse in Asia during the Barremian (Poropat et al., 2016). Recent evidence indicates, however, that the basal somphospondylan Euhelopus could be as early as Berriasian and accordingly could represent the oldest known somphospondylan, suggesting that somphospondylans could have originated in eastern Asia (Xu & Li, 2015).
Titanosauria, defined as the least inclusive clade containing both Andesaurus delgadoi and Saltasaurus loricatus, originated in the Early Cretaceous (Mannion et al., 2013). The oldest known valid titanosaurians include Tapuiasaurus from the Aptian of Brazil (Zaher et al., 2011), Malawisaurus and Karongasaurus from the Aptian of Africa (Gomani, 2005), Daxiatitan and Yongjinglong from the Early Cretaceous of China (You et al., 2008; Li et al., 2014), and Tengrisaurus from the Barremian-Aptian Murtoi Formation of south-central Siberia, Russia (Averianov & Skutschas, 2017). A South American origin has been inferred for Titanosauria using a model-based phylogenetic approach (Gorscak & O’Connor, 2016). However, the analysis in question was carried out before Tengrisaurus, which likely represents the earliest titanosaurian from Asia, was reported. Considering the Asian provenance of Tengrisaurus and the fact that many of the early titanosaurians mentioned above are also from Asia, it seems possible that titanosaurians originated in Asia and subsequently formed endemic groupings in various parts of the world as the continents moved apart during the Cretaceous. It is unclear, however, whether any of the Zhuzhou specimens represent titanosaurians as opposed to basal titanosauriforms.
Some of the Zhuzhou sauropod specimens display an intriguing combination of characters typically seen in basal sauropods, diplodocoids and titanosauriforms. For example, pneumatic fossae and a sharp keel are present on the ventral surface of the probable anterior cervical ZGT002. Similar features are otherwise known mainly in rebbachisaurids and dicraeosaurids (Whitlock, D’Emic & Wilson, 2011; Ibiricu et al., 2013; Tschopp, Mateus & Benson, 2015), although they also occur in the anterior cervical vertebrae of the basal sauropod Shunosaurus (Dong, Zhou & Zhang, 1983). The ischium ZMW148 has a relatively slender posteroventral shaft and an acetabular surface that is narrowest in its central portion, as in the ischia of most diplodocoids (Mannion et al., 2012; Wilson & Allain, 2015). The presence of diplodocoid traits in some of the Zhuzhou specimens suggests that patterns of character evolution in neosauropods were more complex than previously appreciated, reduces the sharpness of the distinction in skeletal anatomy between titanosauriforms and diplodocoids, and reinforces the need for caution in referring incomplete specimens to one clade or the other. In particular, diplodocoids are currently represented in the Asian fossil record only by the recently discovered dicraeosaurid Lingwulong from the Toarcian-Bajocian of Ningxia Autonomous Region in northern China. Various specimens from the Cretaceous of Asia have been previously suggested to also represent diplodocoids, but have been placed in or near Titanosauriformes in recent phylogenetic analyses; this applies, for example, to the holotype of Nemegtosaurus from the Upper Cretaceous of Mongolia (Wilson, 2005), and to an isolated anterior caudal vertebra from the Lower Cretaceous of Shandong Province, China (Upchurch & Mannion, 2009; Whitlock, D’Emic & Wilson, 2011). At present, the absence of diplodocoids from the Cretaceous of Asia is a clear palaeobiogeographic datum, and the occurrence of a few typically diplodocoid characters among the Zhuzhou sample of titanosauriform material implies that future claims of diplodocoid occurrences in the Cretaceous of Asia should be treated with skepticism unless they are based on more substantial evidence than a small number of features occurring in a fragmentary specimen.
Conclusions
This contribution represents the first detailed study of sauropod material from the Upper Cretaceous Daijiaping Formation of Tianyuan District, Zhuzhou City, Hunan Province, southern China. Most of the individual bones are clearly referable to Titanosauriformes, and therefore document a new occurrence of this clade in the Upper Cretaceous of eastern Asia. While none of the specimens are demonstrably of non-titanosauriform origin, some possess features that occur in basal sauropods and diplodocoids, suggesting complex patterns of character evolution within Neosauropoda.